
Det Golgi-apparat, Også kendt som Golgi-kompleks, det er en membrancelleorganel dannet af et sæt flade vesikler stablet sammen; Disse sække har væske indeni. Findes i en bred vifte af eukaryoter, herunder dyr, planter og svampe.
Denne organelle er ansvarlig for forarbejdning, emballering, klassificering, distribution og modifikation af proteiner. Derudover har det også en rolle i syntesen af lipider og kulhydrater. På den anden side forekommer syntesen af komponenterne i cellevæggen i planter i Golgi-apparatet..
Golgi-apparatet blev opdaget i 1888, mens nerveceller blev undersøgt; opdageren, Camillo Golgi, vandt Nobelprisen. Strukturen kan påvises ved sølvkromatfarvning.
Først var organets eksistens tvivlsom for datidens forskere, og de tilskrev Golgi-observationer til enkle artefakter, der er fremstillet af de anvendte teknikker..
Artikelindeks
Golgi-apparatet er en eukaryot organel af membranøs art. Det ligner poser i stakke, selvom organisationen kan variere afhængigt af celletype og organisme. Det er ansvarligt for modifikationen af proteiner efter translation.
For eksempel kan noget kulhydrat tilsættes for at danne et glycoprotein. Dette produkt pakkes og distribueres til cellekammeret, hvor det er nødvendigt, såsom membranen, lysosomer eller vakuoler; det kan også sendes uden for cellen. Det deltager også i syntesen af biomolekyler.
Cytoskeletet (specifikt actin) bestemmer dets placering, og generelt er komplekset placeret i et område af det indre af cellen tæt på kernen og centrosomet.
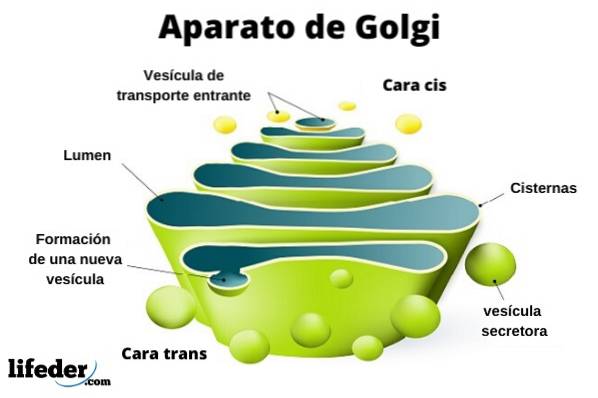
Golgi-komplekset består af et sæt flade, fenestrerede, skiveformede sække, kaldet Golgian cisterner, med variabel tykkelse.
Disse sække er stablet i grupper på fire eller seks cisterner. I en pattedyrcelle kan du finde mellem 40 og 100 batterier forbundet med hinanden.
Golgi-komplekset præsenterer en interessant egenskab: der er polaritet med hensyn til struktur og også funktion.
Du kan skelne mellem cis ansigt og trans ansigt. Den første er relateret til indgangen af proteiner og er placeret nær det endoplasmatiske retikulum. Det andet er udgangsfladen eller produktsekretionen; De er dannet af en eller to tanke, der har en rørform.
Sammen med denne struktur er vesikler, der udgør transportsystemet. Bunkerne med sække er fastgjort til hinanden i en struktur, der minder om en bue eller dato.
Hos pattedyr er Golgi-komplekset fragmenteret i flere vesikler under celledelingsprocesser. Blærerne passerer til dattercellerne og tager igen den traditionelle form af komplekset.
Organiseringen af komplekset er ikke almindelig i alle grupper af organismer. I nogle celletyper er komplekset ikke struktureret som sæt cisterner stablet i grupper; tværtimod er de placeret individuelt. Et eksempel på denne organisation er svampens Saccharomyces cerevisiae.
I nogle encellede organismer, såsom toxoplasma eller trypanosom, tilstedeværelsen af kun en membranbunke er rapporteret.
Alle disse undtagelser indikerer, at stabling af strukturer ikke er afgørende for at udføre deres funktion, skønt nærheden mellem poserne gør transportprocessen meget mere effektiv..
Tilsvarende mangler nogle basale eukaryoter disse cisterner; for eksempel svampe. Dette bevis understøtter teorien om, at apparatet dukkede op i en senere slægt end de første eukaryoter..
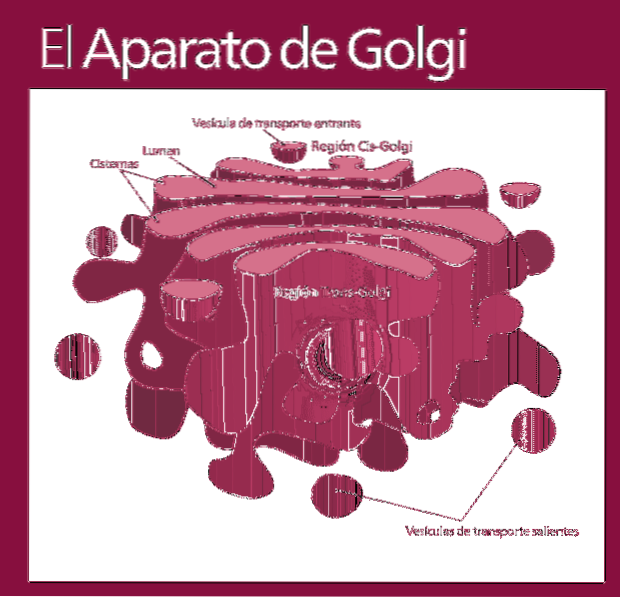
Funktionelt er Golgi-komplekset opdelt i følgende rum: cis-netværket, de stablede sække - som igen er opdelt i mellem- og trans-underrummet - og trans.
Molekylerne, der vil blive modificeret, kommer ind i Golgi-komplekset i samme rækkefølge (cis-netværk efterfulgt af underrum, der endelig udskilles i trans-netværket).
De fleste reaktioner forekommer i de mest aktive zoner: trans- og midterunderrum.
Golgi-komplekset har som hovedfunktion post-translationel modifikation af proteiner takket være de enzymer, de har indeni.
Disse modifikationer inkluderer processer med glycosylering (tilsætning af kulhydrater), phosphorylering (tilsætning af en phosphatgruppe), sulfatering (tilsætning af en phosphatgruppe) og proteolyse (nedbrydning af proteiner).
Desuden er Golgi-komplekset involveret i syntesen af specifikke biomolekyler. Hver af dens funktioner er beskrevet i detaljer nedenfor:
I Golgi-apparatet forekommer modifikationen af et protein til et glycoprotein. Den typiske sure pH i det indre af organellen er afgørende for, at denne proces finder sted normalt..
Der er en konstant udveksling af materialer mellem Golgi-apparatet med det endoplasmatiske retikulum og lysosomerne. I det endoplasmatiske retikulum gennemgår proteiner også ændringer; disse inkluderer tilsætningen af et oligosaccharid.
Når disse molekyler (N-oligosaccharider) kommer ind i Golgi-komplekset, modtager de en række yderligere ændringer. Hvis dette molekyls skæbne skal bæres uden for cellen eller modtages i plasmamembranen, forekommer der særlige ændringer.
Disse modifikationer inkluderer følgende trin: fjernelse af tre mannoserester, tilsætning af N-acetylglucosamin, fjernelse af to mannoser og tilsætning af fucose, yderligere to N-acetylglucosamin-, tre galactose- og tre sialinsyrerester..
I modsætning hertil modificeres de proteiner, der er bestemt til lysosomer, på følgende måde: der er ingen fjernelse af mannoser som et indledende trin; i stedet forekommer phosphorylering af disse rester. Dette trin forekommer i cis-regionen i komplekset.
Dernæst fjernes N-acetylglucosamin-grupperne, hvorved manoserne efterlades med phosphatet tilsat til oligosaccharidet. Disse phosphater indikerer, at proteinet skal målrettes specifikt mod lysosomer..
Receptorerne, der er ansvarlige for at genkende de phosphater, der indikerer deres intracellulære skæbne, er placeret i trans-netværket.
I Golgi-komplekset forekommer syntesen af glycolipider og sphingomyelin ved anvendelse af ceramid (tidligere syntetiseret i det endoplasmatiske retikulum) som oprindelsesmolekyle. Denne proces er i modstrid med den for resten af de phospholipider, der udgør plasmamembranen, som er afledt af glycerol..
Sphingomyelin er en klasse af sfingolipid. Det er en rigelig bestanddel af pattedyrsmembraner, især nerveceller, hvor de er en del af myelinskeden.
Efter deres syntese transporteres de til deres endelige placering: plasmamembranen. Deres polære hoveder er placeret mod ydersiden af celleoverfladen; disse elementer har en specifik rolle i cellegenkendelsesprocesser.
I planteceller bidrager Golgi-apparatet til syntesen af de polysaccharider, der udgør cellevæggen, specifikt hemicellulose og pektiner. Gennem vesikulær transport bæres disse polymerer uden for cellen.
I grøntsager er dette trin afgørende, og ca. 80% af netværksaktiviteten tildeles syntesen af polysaccharider. Faktisk er hundreder af disse organeller blevet rapporteret i planteceller..
De forskellige biomolekyler - proteiner, kulhydrater og lipider - overføres til deres cellulære destinationer af Golgi-komplekset. Proteiner har en slags "kode", der er ansvarlig for at informere den destination, den hører til.
De transporteres i vesikler, der forlader trans-netværket og rejser til det specifikke celleområde.
Proteiner kan føres til membranen ad en specifik konstitutiv vej. Derfor er der en kontinuerlig inkorporering af proteiner og lipider i plasmamembranen. Proteiner, hvis endelige destination er Golgi-komplekset, bevares ved dette.
Ud over den konstitutive vej er andre proteiner bestemt til ydersiden af cellen og forekommer gennem signaler fra miljøet, det være sig hormoner, enzymer eller neurotransmittere..
For eksempel pakkes fordøjelsesenzymer i bugspytkirtelceller i vesikler, der kun udskilles, når tilstedeværelsen af mad opdages..
Nyere forskning rapporterer om eksistensen af alternative veje for membranproteiner, der ikke passerer gennem Golgi-apparatet. Imidlertid disse veje omgå "Ukonventionelle" debatteres i litteraturen.
Der er fem modeller til forklaring af proteinhandel i apparatet. Den første involverer trafik af materiale mellem stabile rum, hver har de nødvendige enzymer til at udføre specifikke funktioner. Den anden model involverer modning af cisternerne gradvist.
Den tredje foreslår også modning af poserne, men med inkorporering af en ny komponent: den rørformede transport. Ifølge modellen er tubuli vigtige i trafikken i begge retninger.
Den fjerde model foreslår, at komplekset fungerer som en enhed. Den femte og sidste model er den nyeste og fastholder, at komplekset er opdelt i forskellige rum.
I visse celletyper har Golgi-komplekset specifikke funktioner. Cellerne i bugspytkirtlen har specialiserede strukturer til udskillelse af insulin.
De forskellige blodtyper hos mennesker er et eksempel på forskellige glycosyleringsmønstre. Dette fænomen forklares ved tilstedeværelsen af forskellige alleler, der koder for glucotransferase.



Endnu ingen kommentarer