Det cytosin Det er en nitrogenbaseret pyrimidintype, der anvendes til biosyntese af cytidin-5'-monophosphat og deoxycytidin-5'-monophosphat. Disse forbindelser tjener til henholdsvis biosyntese af deoxyribonukleinsyre (DNA) og ribonukleinsyre (RNA). DNA lagrer genetisk information, og RNA har forskellige funktioner.
I levende ting findes cytosin ikke i fri form, men danner almindeligvis ribonukleotider eller deoxyribonukleotider. Begge typer forbindelser har en phosphatgruppe, en ribose og en nitrogenbase..
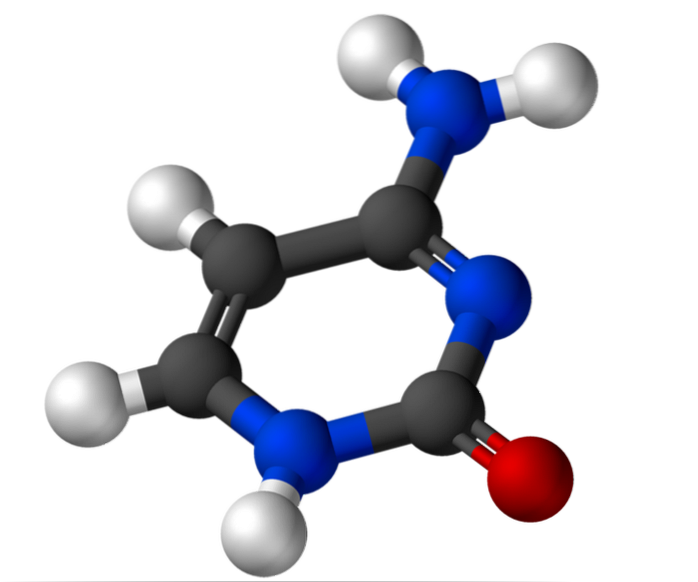
2-carbon af ribose har en hydroxylgruppe (-OH) på ribonukleotider og et hydrogenatom (-H) på deoxyribonukleotider. Afhængig af antallet af tilstedeværende phosphatgrupper er der cytidin-5'-monophosphat (CMP), cytidin-5'-diphosphat (CDP) og cytidin-5'-triphosphat (CTP).
De deoxygenerede ækvivalenter kaldes deoxycytidin-5'-monophosphat (dCMP), deoxycytidin-5'-diphosphat (dCDP) og deoxycytidin-5'-triphosphat (dCTP)..
Cytosin, i dets forskellige former, deltager i forskellige funktioner, såsom DNA- og RNA-biosyntese, glycoproteinbiosyntese og regulering af genekspression..
Artikelindeks
Cytosin, 4-amino-2-hydroxypyrimidin, har den empiriske formel C4H5N3O, hvis molekylvægt er 111,10 g / mol og renses som et hvidt pulver.
Strukturen af cytosin er en plan aromatisk heterocyklisk ring. Bølgelængden med maksimal absorbans (ʎmaks) er ved 260 nm. Cytosins smeltetemperatur overstiger 300 ° C.
For at danne et nukleotid er cytosin bundet kovalent gennem nitrogen 1 via en N-beta-glycosidbinding til 1'-carbonet i ribose. 5'-carbonet forestres med en phosphatgruppe.
Pyrimidinnukleotidbiosyntese har en fælles vej, der består af seks enzymkatalyserede trin. Vejen begynder med carbamoylphosphatbiosyntese. I prokaryoter er der kun et enzym: carbamoylphosphatsyntase. Dette er ansvarlig for syntesen af pyrimidiner og glutamin. I eukaryoter er der carbamoylphosphatsyntase I og II, som henholdsvis er ansvarlige for biosyntese af glutamin og pyrimidiner.
Det andet trin består i dannelsen af N-carbamoylaspartat ud fra carboylphosphat og aspartat, en reaktion katalyseret af aspartattranscabamoylase (ATCase).
Det tredje trin er syntesen af L-dihydrorotat, som forårsager lukningen af pyrimidinringen. Dette trin katalyseres af dihydrootase..
Det fjerde trin er dannelsen af orotat, som er en redoxreaktion katalyseret af dihydroorotatdehydrogenase..
Det femte trin består i dannelsen af orotidylat (OMP) ved anvendelse af phosphoribosylpyrophosphat (PRPP) som substrat og orotatphosphoribosyltransferase som en katalysator..
Det sjette trin er dannelsen af uridylat (uridin-5'-monophosphat, UMP), en reaktion katalyseret af en OMP-decarboxylase..
De næste trin består af phosphorylering af UMP, katalyseret af kinaser, til dannelse af UTP, og overførsel af en aminogruppe fra glutamin til UTP til dannelse af CTP, en reaktion katalyseret af CTP-synthetase..
Hos pattedyr forekommer regulering på niveauet af carbamoylphosphatsyntase II, et enzym der findes i cytosolen, hvorimod carbamoylphosphatsyntase I er mitokondrie.
Carbamoylphosphatsyntase II reguleres af negativ feedback. Dens regulatorer, UTP og PRPP, er henholdsvis hæmmer og aktivator af dette enzym.
I ikke-levervæv er carbamoylphosphatsyntase II den eneste kilde til carbamoylphosphat. Mens det er i leveren, under betingelser med overskydende ammoniak, producerer carbamoylphosphatsyntase I i mitokondrierne carbamoylphosphat, der transporteres til cytosolen, hvorfra det kommer ind i pyrimidinbiosyntesevejen..
Et andet reguleringspunkt er OMP-decarboxylase, som reguleres af konkurrerende hæmning. Dets reaktionsprodukt, UMP, konkurrerer med OMP om bindingsstedet på OMP-decarboxylase..
Genanvendelse af pyrimidiner har den funktion at genbruge pyrimidiner uden behov for de novo biosyntese og undgå den nedbrydende vej. Genbrugsreaktionen katalyseres af pyrimimidinphosphoribosyltransferase. Den generelle reaktion er som følger:
Pyrimidin + PRPP -> pyrimidinnukleosid 5'-monophosphat + PPi
Hos hvirveldyr findes pyrimimidinphosphoribosyltransferase i erythrocytter. Substratpyrimidinerne for dette enzym er uracil, thymin og orotat. Cytosin genvindes indirekte fra uridin-5'-monophosphat.
Under DNA-replikering kopieres informationen i DNA'et til DNA'et af en DNA-polymerase..
RNA-biosyntese kræver deoxynukleotidtriphosphat (dNTP), nemlig: deoxythymidintriphosphat (dTTP), deoxycytidintriphosphat (dCTP), deoxyadenintriphosphat (dATP) og deoxyguanintrifosfat (dGTP). Reaktionen er:
(DNA)n rester + dNTP -> (DNA)n + 1 rest + PPi
Hydrolyse af uorganisk pyrophosphat (PPi) giver energi til RNA-biosyntese.
I DNA-dobbelthelixen er en enstrenget purin forbundet med den modsatstrengede pyrimidin ved hjælp af hydrogenbindinger. Således er cytosin altid bundet til guanin ved hjælp af tre hydrogenbindinger: adenin er bundet til thymin med to hydrogenbindinger..
Hydrogenbindinger brydes, når en opløsning af oprenset nativt DNA ved pH 7 udsættes for temperaturer over 80 ºC. Dette får DNA-dobbelthelixen til at danne to separate tråde. Denne proces er kendt som denaturering..
Temperaturen, ved hvilken 50% DNA denatureres, er kendt som smeltetemperaturen (Tm). DNA-molekyler, hvis forhold mellem guanin og cytosin er højere end thymin og adenin, har højere Tm-værdier end dem, hvis forhold mellem baser er invers.
Det ovenfor beskrevne udgør det eksperimentelle bevis for, at et større antal hydrogenbindinger bedre stabiliserer de native DNA-molekyler..
For nylig blev det fundet, at DNA fra kernen i humane celler kan vedtage interstruktionsmotiv (iM) strukturer. Disse strukturer forekommer i regioner rig på cytosin.
IM-strukturen består af fire DNA-tråde, i modsætning til klassisk dobbeltstrenget DNA, som har to tråde. Mere specifikt er to parallelle duplexkæder afskåret i en antiparallel orientering og holdes sammen af et par hemiprotonerede cytosiner (C: C+).
I det menneskelige genom findes iM-strukturer i regioner som promotorer og telomerer. Antallet af iM-strukturer er højere under G1 / S-fasen af cellecyklussen, hvor transkriptionen er høj. Disse regioner er proteingenkendelsessteder involveret i aktiveringen af transkriptionsmaskineriet..
På den anden side har DNA en tendens til at antage A-helix-formen under dehydratiserende betingelser i regioner, der er rige på på hinanden følgende guaninbasepar (C). Denne form er typisk for RNA og blandede DNA-RNA dobbeltbånd under transkription og replikation og på bestemte tidspunkter, når DNA er bundet til proteiner..
På hinanden følgende baseregioner af cytosin har vist sig at skabe et elektropositivt plaster i den største kløft af DNA. Derfor menes disse regioner at binde sig til proteiner, der disponerer visse genomiske regioner for genetisk skrøbelighed..
Under transkription kopieres informationen i DNA til RNA af en RNA-polymerase. RNA-biosyntese kræver nukleosidtriphosphat (NTP), nemlig: cytidintriphosphat (CTP), uridintriphosphat (UTP), adenintriphosphat (ATP) og guanintriphosphat (GTP). Reaktionen er:
(RNA)n rester + NTP -> (RNA)n + 1 rest + PPi
Hydrolyse af uorganisk pyrophosphat (PPi) giver energi til RNA-biosyntese.
Den sekventielle overførsel af hexoser til dannelse af oligosaccharider, O-bundet til proteiner, sker fra nukleotidforløbere.
Hos hvirveldyr består det sidste trin i O-bundet oligosaccharidbiosyntese af tilsætningen af to sialinsyrerester (N-acetylneuramin) fra en forstadium til cytidin-5'-monophosphat (CMP). Denne reaktion forekommer i trans Golgi sac.
Tetrahydrofolatsyre (FH4) er en kilde til -CH grupper3, og er nødvendig til biosyntese af dTMP fra dUMP. Derudover dannes FH2. Reduktionen af FH2 til FH4 kræver en reduktase af folat og NADPH. Nogle folatreduktasehæmmere, såsom aminopterin og methotrexat, anvendes i kræftbehandlinger.
Methotrexan er en konkurrencedygtig hæmmer. Folatreduktase binder med 100 gange mere affinitet til denne hæmmer end til dets substrat. Aminopterin fungerer på en lignende måde.
Inhiberingen af folatreduktase hindrer indirekte biosyntese af dTMP og derfor af dCTP. Direkte inhibering forekommer af inhibitorer af thymidylatsyntetaseenzymet, som katalyserer dTMP fra dUMP. Disse hæmmere er 5-fluorouracil og 5-fluor-2-deoxyuridin..
For eksempel er 5-fluoracyl ikke i sig selv en hæmmer, men omdannes først i genbrugsvejen til deoxyuridinmphosphat d (FdUMP), som binder og hæmmer thymidylatsyntetase..
Stoffer, der er analoge med glutamin, azaserin og acivicin, hæmmer glutaminamidotransferase. Azarin var et af de første stoffer, der blev opdaget, der fungerer som en selvmordsinaktivator.



Endnu ingen kommentarer