Det sphingomyelin det er den mest forekommende sfingolipid i dyrevæv: dets tilstedeværelse er fundet i alle cellemembraner, der er undersøgt til dato. Det har strukturelle ligheder med phosphatidylcholin med hensyn til den polære hovedgruppe, hvorfor det også er klassificeret som et phospholipid (phosphosphingolipid).
I 1880'erne isolerede videnskabsmand Johann Thudichum en etheropløselig lipidkomponent fra hjernevæv og kaldte den sphingomyelin. Senere, i 1927, blev strukturen af dette sphingolipid rapporteret som N-acyl-sphingosin-1-phosphocholin.
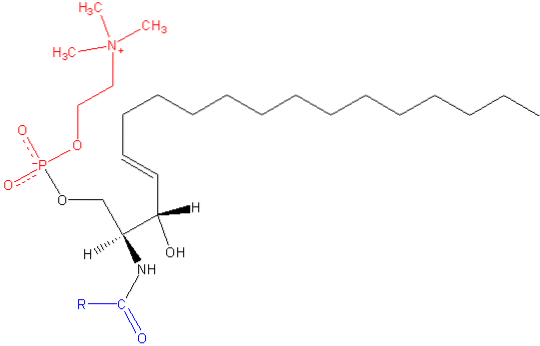
Ligesom de andre sfingolipider har sphingomyelin både strukturelle og celle-signalfunktioner og er især rigeligt i nervevæv, specifikt i myelin, en kappe, der dækker og isolerer axoner fra visse neuroner.
Dens fordeling er blevet undersøgt gennem subcellulær fraktionering og enzymatisk nedbrydningseksperimenter med sfingomyelinaser, og resultaterne indikerer, at mere end halvdelen af sphingomyelin i eukaryote celler findes i plasmamembranen. Dette afhænger dog af celletypen. I fibroblaster udgør det for eksempel næsten 90% af de samlede lipider.
Dysreguleringen af syntesen og metabolismen af dette lipid fører til udviklingen af komplekse patologier eller lipidose. Et eksempel på disse er arvelig Niemann-Pick sygdom, der er karakteriseret ved hepatosplenomegali og progressiv neurologisk dysfunktion.
Artikelindeks
Sphingomyelin er et amfipatisk molekyle sammensat af et polært hoved og to apolære haler. Den polære hovedgruppe er et phosphocholinmolekyle, så det kan se ud som glycerophospholipidphosphatidylcholin (PC). Der er imidlertid væsentlige forskelle med hensyn til grænsefladen og den hydrofobe region mellem disse to molekyler..
Den mest almindelige base i et sphingomyelinmolekyle fra pattedyr er ceramid, der består af sfingosin (1,3-dihydroxy-2-amino-4-octadecen), som har en dobbeltbinding i trans mellem kulstofferne i position 4 og 5 i carbonhydridkæden. Dets mættede derivat, sphinganin, er også almindelig, men findes i mindre grad.
Længden af de hydrofobe haler af sfingomyelin varierer fra 16 til 24 carbonatomer, og fedtsyresammensætningen varierer afhængigt af vævet.
Sfingomyelinerne i den hvide substans i den menneskelige hjerne har f.eks. Nervonsyre, de af det grå stof indeholder hovedsageligt stearinsyre, og den udbredte form i blodplader er arachidonat..
Der er generelt en forskel i længde mellem de to fedtsyrekæder af sfingomyelin, hvilket synes at favorisere "interdigitering" fænomener mellem carbonhydrider i modsatte monolag. Dette giver membranen særlig stabilitet og særlige egenskaber sammenlignet med andre membraner, der er fattigere i dette sfingolipid..
I molekylets grænsefladeområde har sphingomyelin en amidgruppe og en fri hydroxyl ved carbon 3, som kan tjene som donorer og acceptorer af hydrogenbindinger til intra- og intermolekylære bindinger, der er vigtige i definitionen af sidedomæner og interaktion med forskellige. typer af molekyler.
Produkterne af sfingosinmetabolisme -ceramid, sfingosin, sfingosin 1-phosphat og diacylglycerol- er vigtige cellulære effektorer og giver det en rolle i flere cellulære funktioner, såsom apoptose, udvikling og aldring, cellesignalering, blandt andre..
Takket være den tredimensionale "cylindriske" struktur af sfingomyelin kan dette lipid danne mere kompakte og ordnede membrandomæner, hvilket har vigtige funktionelle implikationer fra proteinets synspunkt, da det kan etablere specifikke domæner for nogle integrerede membranproteiner.
Lipidflåder, membranfaser eller ordnede mikrodomæner af sfingolipider, såsom sfingomyelin, nogle glycerophospholipider og cholesterol, repræsenterer stabile platforme til associering af membranproteiner med forskellige funktioner (receptorer, transportører osv.).
Caveolae er invaginationer af plasmamembranen, der rekrutterer proteiner med GPI-ankre og er også rige på sfingomyelin.
Kolesterol på grund af dets strukturelle stivhed påvirker cellemembranernes struktur markant, især i aspekter relateret til fluiditet, hvorfor det betragtes som et væsentligt element.
Fordi sfingomyeliner har både hydrogenbindingsdonorer og -acceptorer, antages det, at de er i stand til at danne mere “stabile” interaktioner med kolesterolmolekyler. Derfor siges det, at der er en positiv sammenhæng mellem niveauet af kolesterol og sfingomyelin i membranerne..
Syntese af sfingomyelin forekommer i Golgi-komplekset, hvor ceramidet, der transporteres fra det endoplasmatiske reticulum (ER), modificeres ved overførsel af et phosphocholinmolekyle fra phosphatidylcholin med den samtidig frigivelse af et diacylglycerolmolekyle. Reaktionen katalyseres af SM-synthase (ceramid: phosphatidylcholin-phosphocholin-transferase).
Der er også en anden vej til sphingomyelinproduktion, der kan forekomme ved at overføre en phosphoethanolamin fra phosphatidylethanolamin (PE) til ceramid med efterfølgende phosphoethanolamin-methylering. Dette menes at være særlig vigtigt i nogle PE-rige nervevæv..
Sphingomyelin-syntase findes på den luminale side af Golgi-kompleksmembranen, hvilket er i overensstemmelse med den ekstra cytoplasmatiske placering af sphingomyelin i de fleste celler.
På grund af egenskaberne ved den polære gruppe af sfingomyelin og det tilsyneladende fravær af specifikke translokaser afhænger den topologiske orientering af dette lipid af enzymsyntasen..
Sphingomyelin-nedbrydning kan forekomme i både plasmamembranen og lysosomer. Lysosomal hydrolyse til ceramid og phosphocholin afhænger af sur sfingomyelinase, et opløseligt lysosomalt glycoprotein, hvis aktivitet har en optimal pH på omkring 4,5.
Hydrolyse i plasmamembranen katalyseres af en sfingomyelinase, der fungerer ved pH 7,4, og som kræver divalent magnesium- eller manganioner til dens drift. Andre enzymer, der er involveret i metabolisme og genanvendelse af sfingomyelin, findes i forskellige organeller, der forbinder hinanden gennem vesikulære transportveje..



Endnu ingen kommentarer