
Det fibronectin Det er en klasse glykoprotein, der hører til den ekstracellulære matrix. Denne type protein er generelt ansvarlig for at fæstne eller binde cellemembranen til de kollagenfibre, der findes på ydersiden.
Navnet "fibronectin" kommer fra et ord, der består af to latinske ord, det første er "fiber" hvilket betyder fiber eller glødetråd og det andet "Necter" hvilket betyder at forbinde, linke, indsætte eller binde.
Fibronectin blev først visualiseret i 1948 som en kontaminant af fibrinogen fremstillet ved Cohns kolde ethanolfraktioneringsproces. Dette blev identificeret som et unikt plasmaglycoprotein, der havde egenskaberne ved et koldt uopløseligt globulin..
Dette protein har en høj molekylvægt og har været forbundet med en lang række funktioner inden i væv. Disse inkluderer adhæsion mellem celle og celle, organisering af cytoskelettet, onkogen transformation, blandt andre..
Fibronectin distribueres til mange steder i kroppen gennem sin opløselige form i blodplasma, cerebrospinalvæske, synovialvæske, fostervand, sædvæske, spyt og inflammatoriske ekssudater..
Forskere har rapporteret, at plasmakoncentrationen af fibronectin stiger, når gravide kvinder lider af præeklampsi. Denne stigning i koncentrationen af fibronectin er således blevet inkorporeret af specialister til at diagnosticere denne tilstand..
Artikelindeks
Fibronectiner er store glycoproteiner, som har en molekylvægt på ca. 440 kDa. De består af ca. 2.300 aminosyrer, som repræsenterer 95% af proteinet, da de øvrige 5% er kulhydrater.
De forskellige analyser, der er udført på den genomiske og transkriptomiske sekvens (messenger RNA) af proteinet, har vist, at det er sammensat af tre blokke af gentagne homologe sekvenser med længder på 45, 60 og 90 aminosyrer hver..
De tre typer af sekvenser udgør mere end 90% af den samlede struktur af fibronectiner. Homologe type I- og II-sekvenser er sløjfer forbundet med hinanden ved hjælp af disulfidbroer. Disse sløjfer indeholder henholdsvis 45 og 60 aminosyrerester..
Homologe type III-sekvenser svarer til 90 aminosyrer arrangeret på en lineær måde og uden interne disulfidbroer. Imidlertid har nogle af de interne aminosyrer i homologe type III-sekvenser frie sulfvalente grupper (R-S-H).
De tre homologe sekvenser foldes og organiseres i en mere eller mindre lineær matrix til dannelse af to "dimere arme" med næsten identiske proteinunderenheder. Forskelle mellem de to underenheder opstår som følge af modningshændelser efter transkription.
Fibronectiner kan generelt ses på to måder. En åben form, der observeres, når de aflejres på overfladen af membranen, og som de er klar til at binde med en anden komponent i celleudvendigheden. Denne form ses kun ved elektronmikroskopi..
Den anden form kan ses i fysiologiske løsninger. Enderne af hver arm eller forlængelse foldes mod proteinets centrum og forbinder sig gennem carboxylenderne på de kollagenbindende steder. I denne form har proteinet et kugleformet udseende.
Multi-adhæsionsegenskaberne for fibronectin stammer fra tilstedeværelsen af forskellige domæner, der har høje affinitetsværdier for forskellige substrater og proteiner..
De "dimeriske arme" kan opdeles i 7 forskellige funktionelle domæner. Disse klassificeres efter det substrat eller domæne, som hver enkelt binder til. For eksempel: Domæne 1 og Domæne 8 er fibrin-proteinbindende domæner.
Domæne 2 har kollagenbindende egenskaber, domæne 6 er en celleadhæsionsregion, det vil sige, det gør det muligt at forankre sig på næsten enhver membran eller ekstern overflade af celler. Funktionerne i domæner 3 og 5 er stadig ukendte i dag.
I domæne 9 er den carboxylende eller C-terminale ende af proteinet placeret. Celleadhæsionsregionerne i domæne 6 har tripeptidet bestående af aminosyresekvensen Arginin-glycin-asparagin (Arg-Gly-Asp).
Dette tripeptid deles af flere proteiner, såsom kollagen og integriner. Det er den mindste struktur, der kræves til genkendelse af plasmamembranen af fibronectiner og integriner..
Fibronectin repræsenterer, når det er i sin kugleform, en opløselig og fri form i blodet. Imidlertid findes den på celleoverflader og i den ekstracellulære matrix i en ”åben”, stiv og uopløselig form..
Nogle af de processer, hvor fibronectins deltagelse skiller sig ud, er celle-til-celle-binding, cellebinding, forbindelse eller vedhæftning til plasma- eller basalmembraner, stabilisering af blodpropper og sårheling..
Celler klæber til et specifikt sted på fibronectin gennem et receptorprotein kendt som "integrin." Dette protein krydser plasmamembranen til det indre af cellen.
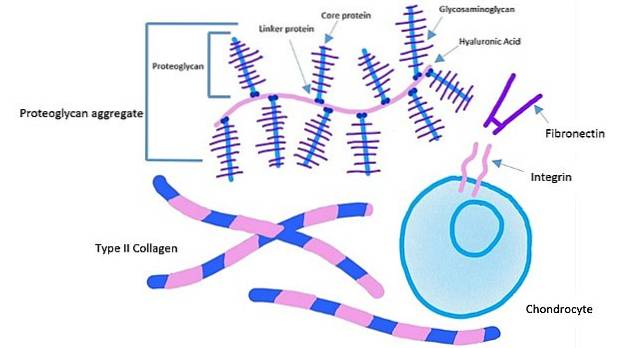
Det ekstracellulære domæne af integriner binder til fibronectin, mens det intracellulære domæne af integriner er bundet til actinfilamenter. Denne type forankring gør det muligt at overføre den spænding, der genereres i den ekstracellulære matrix, til cellernes cytoskelet..
Fibronectiner deltager i sårhelingsprocessen. Disse, i deres opløselige form, afsættes på kollagenfibrene ved siden af såret, hvilket hjælper migrationen af fagocytter, fibroblaster og celleproliferation i det åbne sår..
Den egentlige helingsproces begynder, når fibroblaster "spin" fibronectin netværk. Dette netværk fungerer som en slags stillads eller understøtter aflejring af de nye kollagenfibre, heparansulfat, proteoglycan, chondrotin sultafo og de andre komponenter i den ekstracellulære matrix, der kræves for at reparere vævet..
Fibronectin er også involveret i bevægelsen af epidermale celler, da det gennem det granulære væv hjælper med at omorganisere basalmembranen, der ligger under epidermis i vævene, hvilket hjælper med at keratiniseres..
Alle fibronectiner har vigtige funktioner for alle celler; deltage i processer så forskellige som cellemigration og differentiering, homøostase, sårheling, fagocytose, blandt andre.



Endnu ingen kommentarer