Det heterochromatin Det er en tæt pakket del af kromatin (DNA og histonproteiner) af eukaryote kromosomer. Det er sædvanligvis forbundet med genomets "tavse" regioner, dvs. med dem, der er transkriptionelt inaktive.
Heitz, i 1928, var den første til at skelne mellem to forskellige typer kromatin på eukaryote kromosomer under interfase, der beskriver euchromatin og heterochromatin på basis af deres differentielle komprimering..
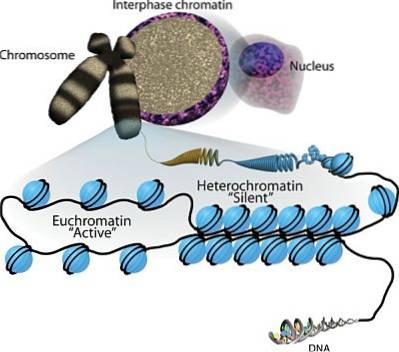
Hvis eukaryote kromosomer farves ved forskellige teknikker, der er specifikke for DNA, afslører mikroskopiske observationer, at der er regioner i disse strukturer, der pletter mere intenst end andre. Disse regioner svarer til de hyperkompakte regioner af heterochromatin.
Heterochromatinization af DNA, det vil sige dets emballage, kan forekomme i en celle som reaktion på forskellige faktorer og kan være fakultativ eller konstituerende.
Konstitutivt heterochromatin er et permanent træk, der generelt nedarves, hvorimod fakultativ heterochromatin måske eller måske ikke er på et kromosom på et givet tidspunkt. Det bedste eksempel på konstituerende heterochromatin er en af de to X-kromosomer hos kvinder.
I eukaryoter "lagrer" og "komprimerer" heterochromatin de store genomer, der karakteriserer dem, især de regioner, der består af gentagne sekvenser, resterende fraktioner af invaderende retro-transposoner, transponerbare elementer, blandt andre..
Artikelindeks
Heterochromatin har ikke en meget anden struktur end kromatin, der er mindre tæt pakket, euchromatin.
At forstå dette er det vigtigt at huske, at eukaryote kromosomer består af et DNA-molekyle, der er forbundet med proteiner kaldet histoner. Otte histoner danner en oktamerisk kerne kendt som et "nukleosom", omkring hvilket DNA ombrydes..
Forbindelsen af DNA med histonproteiner opstår takket være elektrostatiske interaktioner mellem de positive ladninger af de basiske rester af disse proteiner og de negative ladninger af phosphatgrupperne i DNA-strengstrukturen..
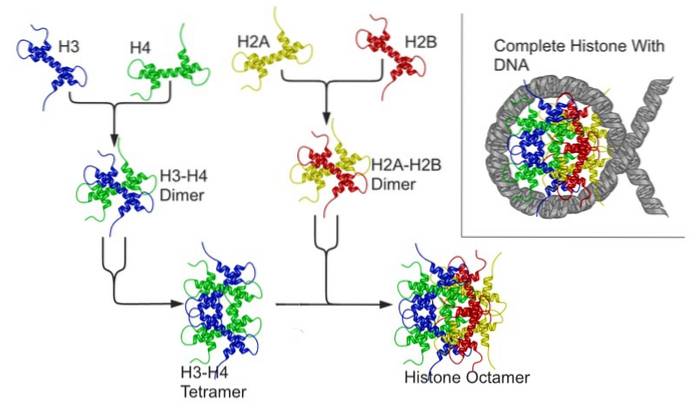
Hver histonoctamer er sammensat af en tetramer af histoner H3 og H4 og af to dimerer af histoner H2A og H2B; omkring hver kerne af histoner rummer ca. 146 basepar DNA.
Nukleosomer "kommer tættere" på hinanden takket være deltagelse af en anden histon kendt som et kryds eller brohiston (linker, på engelsk), som er histon H1.
Kromatin er derefter sammensat af successive nukleosomer, der komprimerer til dannelse af en fibrøs struktur med større tykkelse, men kortere i længden..
Hvert histonprotein er kendetegnet ved tilstedeværelsen af en aminosyre "hale", der kan gennemgå kovalente enzymatiske modifikationer. Disse modifikationer har vist sig at påvirke graden af ekspression eller lyddæmpning af gener associeret med nukleosomer såvel som niveauet af kromatinkomprimering..
Især er heterochromatin karakteriseret ved hypoacetylering af histoner i alle eukaryoter og ved methylering af histon H3 ved lysinrest 9, kun for "højere" eukaryoter..
Enzymerne, der er ansvarlige for at udføre disse modifikationer, er kendt henholdsvis som histondeacetylaser og histonmethyltransferaser..
Udover ændringer i histoner kan DNA også methyleres, hvilket påvirker graden af komprimering af kromatin og svarer til den anden af de to epigenetiske mekanismer til organisering af det eukaryote genom..
Heterochromatin, som kommenteret i begyndelsen, kan være konstitutiv eller fakultativ.
Konstitutivt heterochromatin er særlig rigeligt i genomiske regioner, der har en høj tæthed af gentagne sekvenser (såsom f.eks. Satellitelementer), hvor der er rigelige dæmpede transponerbare elementer i de centromere regioner og i telomerer.
Det siges at være konstitutivt, fordi disse regioner i genomet forbliver kondenserede eller kompakte under celledeling. I en ikke-delende celle er derimod det meste af DNA eukromatisk, og der er kun et par veldefinerede regioner med konstitutivt heterochromatin.
Fakultativ heterochromatin er det, der findes i loci, der er reguleret i de forskellige udviklingsstadier; så det repræsenterer faktisk de "forbigående kondenserede" regioner, der kan ændre sig i henhold til cellulære signaler og genetisk aktivitet.
Da heterochromatin er en vigtig del af de telomere og centromere regioner, udfører det transcendentale funktioner fra celledelingens synspunkt og beskyttelsen af kromosomale ender..
Centromerer fungerer aktivt under celledeling, så duplikatkromosomer bevæger sig mod begge poler i den delende celle, mens resten af generne forbliver inaktive og kompakte..
Komprimering af specifikke regioner af eukaryote kromosomer er synonymt med genetisk dæmpning, da det faktum, at heterochromatin er tæt pakket, indebærer, at transkriptionsmaskineriet ikke er tilgængeligt for de underliggende gensekvenser.
For så vidt angår rekombination undertrykker heterochromatin denne proces og beskytter genomets integritet ved at forbyde "ulovlig" rekombination mellem gentagne DNA-sekvenser spredt gennem genomet. Dette er især vigtigt for bekæmpelsen af "parasitiske" transponerbare elementer, der tavs ved heterochromatinisering..
Indtil for et par år siden troede man, at heterokromatisk DNA var en slags "junk-DNA", da forskere ikke fandt en specifik funktion for sekvenserne inkluderet i disse regioner; husk at mere end 80% af det genomiske DNA fra et menneske f.eks. ikke koder for cellulære proteiner eller for RNA-molekyler med regulerende funktioner.
Det er imidlertid nu kendt, at dannelsen af fakultativ heterochromatisk DNA er af største betydning for reguleringen af mange processer under udvikling og vækst af levende væsener, og at konstituerende heterochromatinregioner spiller en grundlæggende rolle set fra et strukturelt synspunkt. af synspunkt.
Det er blevet foreslået af mange forfattere, at heterochromatin kan have strukturelle funktioner på eukaryote kromosomer. Denne påstand er baseret på det faktum, at heterokromatiske regioner i et givet kromosom adskiller dele af det, der har forskellige mønstre af genetisk "aktivitet".
Med andre ord tjener de heterokromatiske regioner som "afstandsstykker" mellem forskellige transkriptionelt aktive regioner, hvilket kan være af stor betydning ud fra transskriptionen af de gener, der er placeret der..



Endnu ingen kommentarer