
Det humoristisk immunitet, også kendt som det antistofmedierede immunrespons, det er en af de vigtigste forsvarsmekanismer for organismer mod invasion af mikroorganismer eller ekstracellulære toksiner.
Specifikt henviser humoral immunitet til immunitet medieret af blodfaktorer, som er serumproteiner kendt som "antistoffer", der fungerer som respons på infektioner og produceres specifikt som reaktion på tilstedeværelsen af "antigener.".
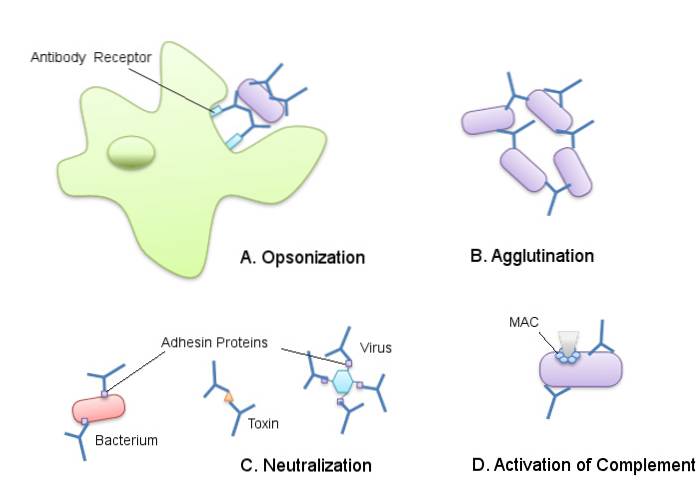
Immunsystemet hos et pattedyr kan opdeles i medfødt immunsystem og adaptivt immunsystem. Det medfødte immunsystem består af de forskellige elementer, der fungerer som fysiske og kemiske barrierer mod indtrængen af invaderende stoffer i kroppen..
Blandt sådanne barrierer er epitel og nogle af de stoffer, der produceres af dem; nogle specifikke celletyper er også involveret, som tilsammen repræsenterer kroppens første forsvarssystem.
Det adaptive eller specifikke immunsystem er lidt mere komplekst og "udviklet", da det udløses som reaktion på eksponering for smitsomme stoffer eller ved kontakt med visse mikroorganismer, selvom begge systemer normalt fungerer sammen.
Det siges at være et specifikt system, fordi det forekommer som reaktion på definerede determinanter og medieres af højt specialiserede celler, der også har evnen til at "huske" og reagere hurtigere og med mere "styrke" eller "effektivitet" på gentagne eksponeringer for samme invaderende agent.
Humoral immunitet er en af underkategorierne for adaptiv eller specifik immunitet, som også klassificeres under cellulær immunitet. Begge typer svar adskiller sig fra hinanden afhængigt af den involverede immunsystemkomponent..
Artikelindeks
Teorien om humoral immunitet, som var et produkt af intense års forskning og debat, foreslår, at immunitet formidles af stoffer, der findes i kropsvæsker eller "humorer"..
Denne teori blev udviklet af mange forskere, der uafhængigt studerede og beskrev mange af de effektorer, der var involveret i sådanne reaktionsmekanismer..
Paul Ehrlich var måske en af de mest indflydelsesrige, da han var den, der gennemførte de mest dybtgående undersøgelser af antigen-antistof-komplementaritet i begyndelsen af 1900'erne..
Den berømte immunolog Rudolph Virchow, i 1858, fastslog, at alle kropslige patologier skyldtes funktionsfejl hos de cellulære elementer, der var ansvarlige for beskyttelsen, snarere end en "uoverensstemmelse mellem de opløselige humorer".
Lidt mere end 25 år senere, i 1884, bragte Eli Metchnikoff den første offentliggørelse af den fagocytiske teori, som i dag former og understøtter de vigtigste baser i teorien om cellemedieret immunitet (cellulær immunitet)..
Mange modstandere af Metchnikoff forsøgte at "modbevise" hans påstande, og det var i 1888, at George Nuttall, der gennemførte en række eksperimenter designet til at teste Metchnikoffs teorier, observerede, at serum fra normale dyr havde en "naturlig toksicitet" mod visse mikroorganismer..
På denne måde blev det populært i den videnskabelige verden, at cellefrie væsker fra "sunde" eller "specielt immuniserede" dyr kunne dræbe bakterier, så det ikke var nødvendigt at ty til celleteori for at forklare medfødt og erhvervet immunitet..
De første, der eksperimentelt bekræftede eksistensen af et humoralt immunrespons, var Emil von Behring og Shibasaburo Kitasato i slutningen af 1800-tallet. Von Behring og Kitasato demonstrerede, at immunresponsen udløst af difteri og stivkrampe skyldtes tilstedeværelsen af antistoffer mod eksotoksin.
I begyndelsen af 1900'erne indså Karl Landsteiner og andre forskere, at andre toksiner og stoffer af ikke-bakteriel oprindelse kunne producere humoristisk immunitet..
Udtrykket "antistof" blev kort efter opfundet som en generalitet for at henvise til de specifikke stoffer, der kunne fungere som antitoksiner mod "antigener".
Ordet antigen var udtrykket brugt til at definere de stoffer, der udløser produktionen af humorale antistoffer.
Både humorale immunresponser og cellulære immunsvar medieres af en type celle kendt som lymfocytter..
De vigtigste hovedpersoner i cellulær immunitet er T-lymfocytter, mens det er B-lymfocytter, der reagerer på tilstedeværelsen af fremmede antigener og bliver de antistofproducerende celler, der er karakteristiske for humoral immunitet..
Humoral immunitet er den vigtigste forsvarsmekanisme mod ekstracellulære mikroorganismer og andre toksiner, mens cellulær immunitet bidrager til eliminering af intracellulære patogener, som er "utilgængelige" til genkendelse af antistoffer..
Ud over det cellulære immunrespons kan det humorale respons opdeles i tre faser: en med genkendelse, en anden med aktivering og en anden med effekt..
Genkendelsesfasen består af binding af antigener til specifikke membranreceptorer på celleoverfladen af modne B-lymfocytter..
Antistoffer fungerer som sådanne "receptorer" og er i stand til at genkende proteiner, polysaccharider, lipider og andre "fremmede" ekstracellulære stoffer..
Aktiveringsfasen begynder med proliferation af lymfocytter efter genkendelse af antigener og fortsætter med differentiering, enten i andre effektorceller, der er i stand til at eliminere antigener, eller i hukommelsesceller, der er i stand til at inducere hurtigere reaktioner efter en ny eksponering for det..
Under effektorfasen er lymfocytterne, der udøver antigenelimineringsfunktioner, kendt som "effektorceller", selvom andre celler normalt deltager, som også deltager i det medfødte immunrespons, og som phagocytose og eliminerer fremmede stoffer.
Antistofferne produceret af lymfocytter eller B-celler har den fysiologiske funktion at neutralisere og eliminere det antigen, der inducerede deres dannelse, og det humorale immunsystem kan reagere på en mangfoldighed af forskellige antigener..
B-lymfocytter stammer fra knoglemarven som reaktion på et defineret antigen (de er specifikke), og dette sker før antigen stimulering. Ekspression af specifikke antistoffer udløser proliferations- og differentieringsreaktioner af mere antistofsekreterende B-celler.
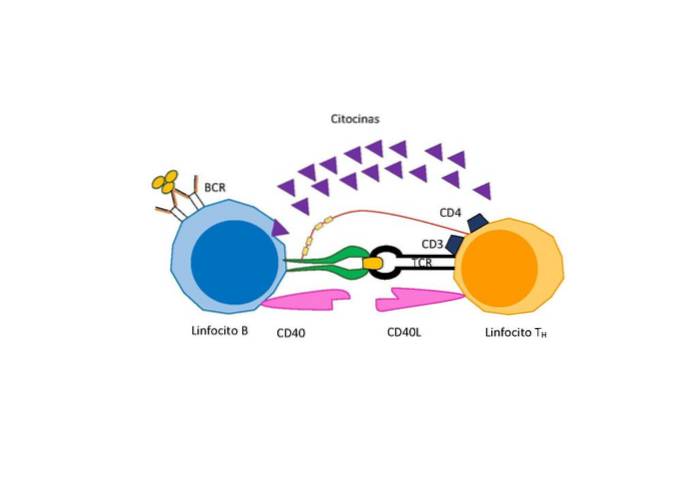
Afhængigt af antigenets art er der imidlertid behov for et yderligere signal til differentiering og proliferation, der gives af en speciel type T-lymfocyt kaldet “hjælper T-lymfocyt”, der udskiller aktiverende faktorer for B-celler..
Da de hovedsageligt findes i blodvæsker, kaldes antistofferne produceret af B-celler immunglobuliner. Disse proteinmolekyler har to tunge og to lette glykoproteinkæder, der er bundet sammen gennem disulfidbroer (SS)..
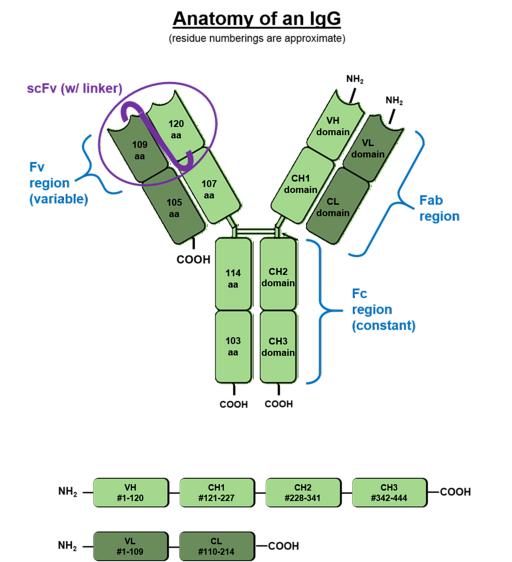
De lette kæder er kendt som "kappa" og "lambda", men der er 5 typer tunge kæder, der er blevet kaldt gamma (G), mu (M), alfa (A), delta (D) og epsilon (E).
Kombinationen af lette og tunge kæder slutter med dannelsen af immunglobulinerne IgG, IgM, IgA, IgD og IgE. Det mest forekommende antistof i serum fra pattedyr over for immunglobulin IgG (ca. 70%).
Hver kæde af et antistof har en aminoterminal og en carboxylterminalende. Den del, der er i stand til at binde til antigener, er ved den aminoterminale ende, men den carboxylterminale region er det, der bestemmer biologisk aktivitet.
Den carboxylterminale region af IgG-antistoffer genkendes specifikt af fagocytiske celler, såsom neutrofiler og makrofager, som har specielle receptorer til det..
Denne genkendelse indebærer kontakt mellem receptoren og antistoffet, og det er denne forening, der letter fagocytose og nedbrydning af antigener inde i fagocytiske celler..
I modsætning til IgG findes de andre klasser af immunglobuliner ikke i sekreter og væv. De er dog lige så nyttige til at fremkalde immunrespons..
IgM-immunglobuliner (10% af serumimmunoglobuliner) er potente aktivatorer af komplementsystemet, derfor fungerer de ved lysering af antigener og øger resistens.
IgA-immunglobuliner (20% af serumimmunglobuliner) produceres i lymfoide væv og behandles og transporteres til lungeslimhinden og mave-tarmkanalen. Disse arbejder i neutralisering af vira og andre antigener, der kommer ind gennem slimhindeoverflader..
IgD er bundet til B-lymfocytter og fungerer som en antigenreceptor, mens IgE (kendt som det allergiske antistof) er bundet til overfladen af mastceller og basofiler gennem specifikke receptorer. Begge immunglobuliner er i meget lav koncentration i serum.
Antistofferne produceret af de vigtigste effektorer af det humorale immunrespons (B-lymfocytter) har evnen til at "inducere" eller "aktivere" forskellige reaktionsmekanismer mod forskellige typer trusler..
F.eks. Er IgG-immunglobuliner aktivatorer af det, der er kendt som "komplementkaskade", som arbejder for at neutralisere virale partikler og derved forhindre deres binding til værtsceller..
Under graviditeten overfører moderen antistoffer til fosteret gennem placenta's trofoblastiske celler, som har receptorer med høj affinitet for carboxylterminen af immunglobuliner, såsom IgG.
Den humorale reaktion på bakterier, der har "kapsler" sammensat af polysaccharider, medieres af immunglobulin M, som fremmer fagocytose af disse mikroorganismer..
Et andet vigtigt eksempel på humoral immunitet er den systemiske reaktion på parasitter, hvor IgE "styrer" deres destruktion gennem eosinofile celler..



Endnu ingen kommentarer