EN polysom det er en gruppe af ribosomer, der er rekrutteret til translation af det samme messenger-RNA (mRNA). Strukturen er bedre kendt som polyribosomet eller det mindre almindelige ergosom..
Polysomer tillader øget produktion af proteiner fra de budbringere, der udsættes for samtidig translation af flere ribosomer. Polysomer deltager også i co-translationelle foldningsprocesser og i erhvervelsen af kvaternære strukturer af nysyntetiserede proteiner..
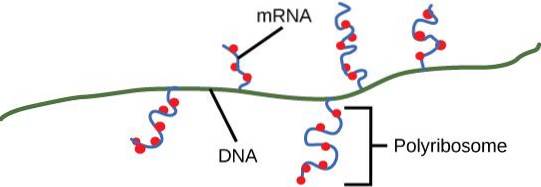
Polysomer sammen med såkaldte P-legemer og stressgranulater styrer skæbnen og funktionen af budbringere i eukaryote celler..
Polysomer er blevet observeret i både prokaryote og eukaryote celler. Dette betyder, at denne type makromolekylær dannelse har en lang historie i den cellulære verden. Et polysom kan bestå af mindst to ribosomer på den samme messenger, men generelt er der mere end to.
I mindst en pattedyrcelle kan der eksistere op til 10.000.000 ribosomer. Mange er blevet observeret at være frie, men en stor del er forbundet med de velkendte polysomer.
Artikelindeks
Ribosomerne i alle levende ting består af to underenheder: den lille underenhed og den store underenhed. Den lille underenhed af ribosomer er ansvarlig for læsning af messenger RNA.
Den store underenhed er ansvarlig for den lineære tilsætning af aminosyrer til det spirende peptid. En aktiv translationel enhed er en, hvor et mRNA har været i stand til at rekruttere og tillade samling af ribosomet. Derefter fortsætter aflæsningen af trillinger i messenger og interaktionen med det tilsvarende belastede tRNA sekventielt..
Ribosomer er polysomernes byggesten. Faktisk kan begge måder at oversætte en messenger sameksistere i den samme celle. Hvis alle komponenter, der udgør cellens translationelle maskineri, renses, ville vi finde fire hovedfraktioner:
I eukaryote celler eksporteres mRNA'er fra kernen som messenger ribonukleoproteiner. Det vil sige, messenger er koblet med flere proteiner, der bestemmer dets eksport, mobilisering og oversættelse..
Blandt dem er der flere, der interagerer med PABP-proteinet bundet til messingens polyA 3 'hale. Andre, såsom CBP20 / CBP80-komplekset, vil binde til 5'-hætte af mRNA.
Frigivelse af CBP20 / CBP80-komplekset og rekruttering af ribosomale underenheder på 5'-hætten definerer dannelsen af ribosomet.
Oversættelse begynder, og nye ribosomer samles på 5'-hætten. Dette sker i et begrænset antal gange, der afhænger af hver messenger og typen af polysom..
Efter dette trin interagerer oversættelsesforlængelsesfaktorer forbundet med hætten i 5'-enden med PABP-proteinet bundet til 3'-enden af mRNA'et. En cirkel dannes således defineret af foreningen af messengerens ikke-oversættelige regioner. Således rekrutteres så mange ribosomer som længden af budbringeren, og andre faktorer tillader det.

Andre polysomer kan vedtage en lineær dobbelt række eller spiralkonfiguration med fire ribosomer pr. Omdrejning. Den cirkulære form har været stærkest forbundet med frie polysomer.
Polysomer dannes på aktive translationelle enheder (oprindeligt monosomer) med sekventiel tilsætning af andre ribosomer på det samme mRNA.
Afhængigt af deres subcellulære placering finder vi tre forskellige typer polysomer, der hver har sine egne særlige funktioner..
De findes frie i cytoplasmaet uden tilsyneladende tilknytning til andre strukturer. Disse polysomer oversætter de mRNA'er, der koder for cytosoliske proteiner.
Da atomhylsteret er en forlængelse af det endoplasmatiske retikulum, kan denne type polysom også være forbundet med den ydre nukleare hylster.
I disse polysomer oversættes mRNA'erne, der koder for to vigtige proteingrupper. Nogle, som er en strukturel del af det endoplasmatiske retikulum eller Golgi-komplekset. Andre, som skal ændres posttranslationsmæssigt og / eller flyttes intracellulært af disse organeller.
Cytoskeletal-associerede polysomer oversætter proteiner fra mRNA'er, der er asymmetrisk koncentreret i visse subcellulære rum.
Det vil sige, at når nogen forlader kernen, mobiliseres nogle messenger-ribonukleoproteiner til det sted, hvor det produkt, de koder, er påkrævet. Denne mobilisering udføres af cytoskelet med deltagelse af proteiner, der binder til polyA-halen af mRNA'et..
Med andre ord distribuerer cytoskelet budbringere efter destination. Denne skæbne er angivet ved proteinets funktion og ved det sted, hvor det skal opholde sig eller handle..
Selv hvis et mRNA transkriberes, betyder det ikke nødvendigvis, at det skal oversættes. Hvis dette mRNA specifikt nedbrydes i cellecytoplasmaet, siges ekspressionen af dets gen at være posttranskriptionelt reguleret..
Der er mange måder at opnå dette på, og en af dem er gennem virkningen af de såkaldte MIR-gener. Slutproduktet af transkriptionen af et MIR-gen er et microRNA (miRNA).
Disse er komplementære eller delvist komplementære til andre budbringere, hvis oversættelse de regulerer (post-transkriptionel dæmpning). Dæmpning kan også involvere specifik nedbrydning af en bestemt messenger.
Alt relateret til translation, dets opdeling, regulering og post-transkriptionel gendæmpning kontrolleres af polysomer..
For at gøre dette interagerer de med andre molekylære makrostrukturer i cellen kendt som P-legemer og stressgranuler. Disse tre kroppe, mRNA'er og microRNA'er, definerer således det proteom, der er til stede i en celle på et givet tidspunkt..



Endnu ingen kommentarer