Det profase det er den første fase af celledeling ved mitose og meiose. Det er fasen efter DNA-syntesetrinnet (S-fase af cellecyklussen). På dette stadium når kromosomerne en høj grad af kondens og individualitet.
I meiose er der to protaser, som er meget forskellige fra hinanden og fra mitose. Kun i meiotisk profase I forekommer for eksempel rekombination. Denne fase er opdelt i forskellige faser: leptoten, zygoten, pachyten, diploten og diakinesis..

Under profasen finder foruden kondensering, der opnås ved de duplikerede kromosomer, forandringsprocesser sted på det ekstra nukleare niveau. Den vigtigste cytoplasmatiske hændelse under profase er dannelsen af den akromatiske spindel ved hver cellepol. Dette gør det muligt at mobilisere kromosomerne i successive faser af celledeling for at sikre deres korrekte segregering..
Der er vigtige forskelle mellem celledelinger i dyreceller og planteceller. Nogle vil blive nævnt senere. I alt er der imidlertid en fuldstændig reorganisering af cellen.
Derfor fokuserer mitose og meiose på skæbnen til DNA og kernen. Men sandheden er, at når en celle deler sig, deler den alt, og alt deltager i processen.
Således gennemgår alle cellulære komponenter radikale ændringer under protosen af mitose og meiose. Det endoplasmatiske retikulum og Golgi-komplekset ser ud til at forsvinde: de ændrer dog kun deres struktur. Mitokondrier og kloroplaster deler sig også, hvilket giver anledning til nye organeller..
Artikelindeks

Dyreceller har en enkelt centriole. Efter afslutning af DNA-syntese som forberedelse til mitose forbereder centriolen sig også til at dele sig.
Centrioler består af et par identiske strukturer kaldet diplosomer, vinkelret på hinanden. Disse adskiller sig, og hver enkelt vil være en form til dannelsen af en ny. Syntesen af det nye diplosom sker, når hvert gammelt diplosom migrerer til modsatte poler i cellen..
Den anden definerende begivenhed af profase, og en, der deles med planteceller, er kromatinkomprimering. Dette er måske det mest bemærkelsesværdige cytologiske element i profase under celledeling..
DNA når en høj grad af komprimering, og for første gang observeres det som et morfologisk individualiseret kromosom.
De komprimerede kromosomer involverer søsterkromatiderne for hver af dem, stadig forenet af den samme centromer. Selvom denne centromer virkelig er dobbelt, opfører den sig som en enkel.
Kromosomerne ses som et X, fordi de er to kopikromatider fastgjort til det samme centrum. Derfor vil hver celle i profase have et dobbelt antal kromatider sammenlignet med et antal centromerer svarende til antallet '2n' af arten..
Det vil sige, at en profase mitotisk celle er diploid efter antal centromerer, men tetraploid (4n) efter antal kromatider..
I planteceller er der en fase forud for profase kaldet preprophase. Som forberedelse til celledeling nedbrydes den store cellevakuol.
Takket være dette dannes et frit eller ledigt cytoplasmatisk bånd kaldet et fragmosom. Dette gør det muligt for plantecellekernen at positionere sig mod cellens ækvator.
Derudover kollapser den kortikale organisering af mikrotubuli mod det samme sted. Dette vil give anledning til det, der er kendt som præ-profasebåndet (BPP)..

Prefase-båndet vises først som en ring, men ender med at dække kernen. Det vil sige at mikrotubuli, der fører cellemembranen internt, alle vil mobilisere mod fragmosomet.
Derefter vil preprophase-båndet, der omgiver ækvatorialkernen, tillade at organisere lokalt det sted, hvor fragmoplasten, der erstatter den, til sidst vises..
Dynamisk set vil mikrotubuli i plantecellen passere fra en fase til en anden uden åbenlyse overgange. Det vil sige fra kortikal arrangement til fragmosom og derfra til fragmoplast.
Stedet for alle disse strukturelle ændringer i plantecellen er det samme, hvor aflejringen af cellepladen vil forekomme. Og derfor repræsenterer det planet, hvor cellen deler sig.
For alt andet er planteprofasen identisk med den, der observeres i profasen om dyreceller.
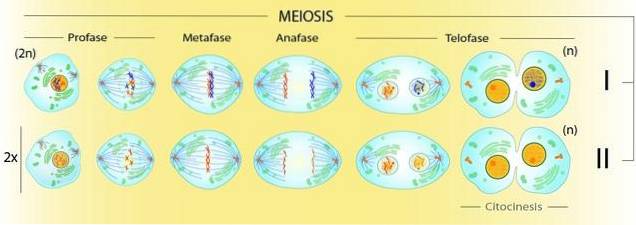
Kun i Mease-fase I forekommer genetisk rekombination. Derfor kræver dannelsen af komplekse strukturer mellem kromosomer, at der er to divisioner i meiose.
Med tidligere DNA-syntese blev søsterkromatiderne produceret på hvert kromosom. Med deres komprimering har vi dobbeltkromosomer, som i meiose derudover parrer mellem homologer.
Dette fører til dannelsen af bivalenter (to interagerende homologe kromosomer). Da hver enkelt er duplikeret, taler vi faktisk om tetrader. Det vil sige af kromatidtetrader forenet i en struktur, der skal løses ved hjælp af to celledelinger.
I det første vil de homologe kromosomer blive adskilt, mens i det andet skal søsterkromatiderne adskilles.
I meiotisk profase I er søsterkromatider organiseret på kompakte proteinstrukturer, der udgør den centrale kromosomakse..
På denne akse dannes det synaptonemiske kompleks (CS), som holder de homologe kromosomer parret sammen. Under profase I vil det synaptonemiske kompleks tillade homologe kromosomer at komme ind i synapser..
I disse faser kan der dannes krydsningspunkter, der er synlige som chiasmas, hvor den genetiske rekombinationsproces vil finde sted. Det vil sige den fysiske udveksling mellem de deltagende DNA-molekyler, der definerer pachytenen.
Før fase II er der ikke forud for tidligere DNA-syntese. Her blev dobbelt kromosomer forbundet med den samme centromer (dobbelt) arvet. Dette skyldes, at DNA-syntese, både i mitose og meiose, kun forekommer i S (syntese) -fasen af cellecyklussen..
I denne anden division har vi fire meiocytter. En meiocyt er en celle, der er et produkt af en meiotisk division.
Profase II vil derfor være ansvarlig for adskillelsen af søsterkromatider fra kromosomer arvet fra profase I. Derfor vil hver meiocyt i slutningen af den meiotiske proces have det haploide sæt af kromosomer af arten.


Endnu ingen kommentarer