Det DNA-emballage er et udtryk, der definerer den kontrollerede komprimering af DNA i cellen. I ingen celle (og faktisk ikke engang i vira) er DNA-fri, løs og i ægte opløsning.
DNA er et ekstremt langt molekyle, der også altid interagerer med et stort udvalg af forskellige proteiner. Til behandling, nedarvning og kontrol af ekspressionen af de gener, den bærer, vedtager DNA en bestemt rumlig organisation. Dette opnås af cellen ved nøje at kontrollere hvert trin i DNA-emballering på forskellige komprimeringsniveauer..
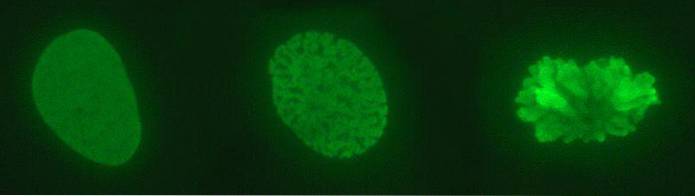
Virus har forskellige emballeringsstrategier for deres nukleinsyrer. En af favoritterne er dannelsen af kompakte spiraler. Man kan sige, at vira er nukleinsyrer pakket i deres egne proteiner, der dækker, beskytter og mobiliserer dem.
I prokaryoter er DNA forbundet med proteiner, der bestemmer dannelsen af komplekse sløjfer i en struktur kaldet nukleoid. Det maksimale niveau af DNA-komprimering i en eukaryot celle er derimod mitotisk eller meiotisk kromosom..
Det eneste tilfælde, hvor et B-DNA ikke er pakket, er et forskningslaboratorium, der forfølger dette formål..
Artikelindeks
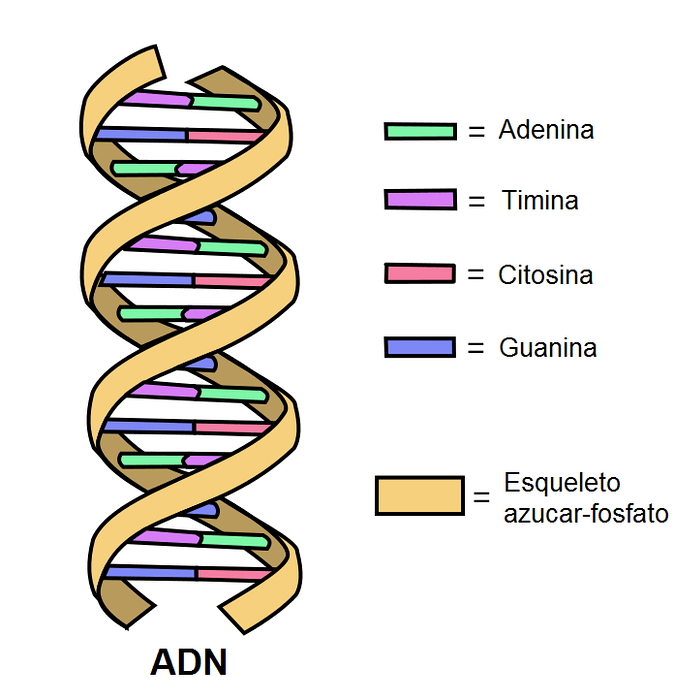
DNA består af to antiparallelle bånd, der danner en dobbelt helix. Hver af dem har et phosphodiesterbindingsskelet, hvorpå sukker, der er bundet til nitrogenholdige baser, er bundet..
Inde i molekylet danner de nitrogenholdige baser i et bånd hydrogenbindinger (to eller tre) med det komplementære bånd.
I et molekyle som dette viser de fleste af de vigtige bindingsvinkler fri rotation. Nitrogen base-sukker, sukker-phosphat gruppe og phosphodiester bindinger er fleksible.
Dette gør det muligt for DNA, set som en fleksibel stang, at vise en vis evne til at bøje og vride. Denne fleksibilitet giver DNA mulighed for at vedtage komplekse lokale strukturer og danne interaktionssløjfer på korte, mellemstore og lange afstande..
Denne fleksibilitet forklarer også, hvordan 2 meter DNA kan opretholdes i hver diploide celle hos et menneske. I en gamet (haplooid celle) ville det være en meter DNA.
Selvom det ikke er en ubrydelig regel, eksisterer bakteriekromosomet som et enkelt supercoiled dobbeltbånds-DNA-molekyle..
Den dobbelte helix vrides mere på sig selv (mere end 10 bp pr. Omdrejning), hvilket giver en vis komprimering. Lokale knuder genereres også takket være manipulationer, der styres enzymatisk.
Derudover er der sekvenser i DNA, der tillader domæner at dannes i store sløjfer. Vi kalder strukturen som følge af superspolen og bestilte sløjfer en nukleoid..
Disse gennemgår dynamiske ændringer takket være nogle proteiner, der giver en vis strukturel stabilitet til det komprimerede kromosom. Graden af komprimering i bakterier og arkæer er så effektiv, at der kan være mere end et kromosom pr. Nukleoid.
Nukleoid komprimerer det prokaryote DNA mindst 1000 gange. Den meget topologiske struktur af nukleoid er en grundlæggende del af reguleringen af de gener, der bæres af kromosomet. Det vil sige, struktur og funktion udgør den samme enhed..
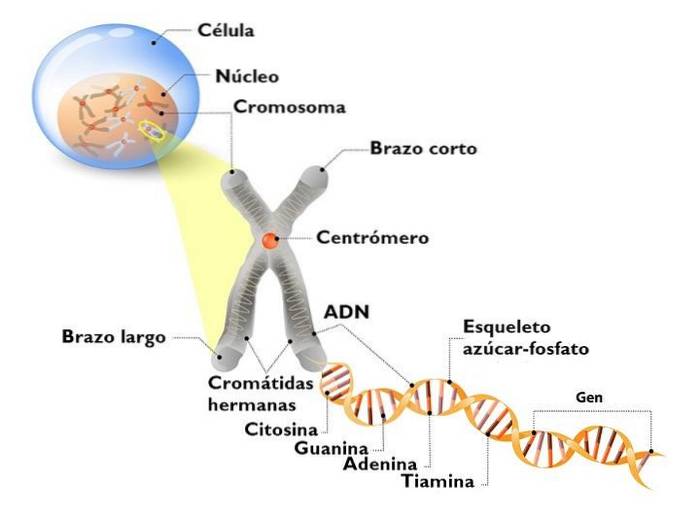
DNA'et i den eukaryote kerne er ikke nøgen. Det interagerer med mange proteiner, hvoraf de vigtigste er histoner. Histoner er små, positivt ladede proteiner, der binder til DNA ikke-specifikt.
I kernen ser vi et komplekst DNA: histoner, som vi kalder kromatin. Det stærkt kondenserede kromatin, som generelt ikke udtrykkes, er heterochromatin. I modsætning hertil er den mindst komprimerede (løsere) eller euchromatin, kromatin med gener, der udtrykkes.
Kromatin har forskellige komprimeringsniveauer. Det mest elementære er nukleosomets; Det efterfølges af magnetfibre og mellemfasekromatinløkker. Det er først, når et kromosom deler sig, at maksimale komprimeringsniveauer vises..
Nukleosomet er den basale organisationsenhed for kromatin. Hvert nukleosom består af en oktamer af histoner, der danner en slags tromme..
Oktameren består af to kopier af hver af histonerne H2A, H2B, H3 og H4. Omkring dem går DNA'et omkring 1,7 gange. Det efterfølges af en brøkdel af frit DNA kaldet en 20 bp linker associeret med histon H1 og derefter et andet nukleosom. Mængden af DNA i et nukleosom og det, der binder det til et andet, er ca. 166 basepar.
Dette DNA-emballeringstrin komprimerer molekylet ca. 7 gange. Det vil sige, vi går fra en meter til lidt over 14 cm DNA.
Denne pakning er mulig, fordi de positive histoner annullerer den negative ladning af DNA'et og den deraf følgende elektrostatiske selvafstødning. Den anden årsag er, at DNA kan foldes på en sådan måde, at det kan vende octamer af histoner..
Fiberen af perler i en halskæde dannet af mange successive nukleosomer vikles yderligere ind i en mere komprimeret struktur.
Selvom vi ikke er klar over, hvilken struktur den faktisk vedtager, ved vi, at den når en tykkelse på ca. 30 nm. Dette er den såkaldte 30 nm fiber; Histon H1 er afgørende for dets dannelse og stabilitet.
30 nm fiberen er den grundlæggende strukturelle enhed af heterochromatin. Det for slappe nukleosomer, det for euchromatin.
30 nm fiberen er dog ikke helt lineær. Tværtimod danner den sløjfer på ca. 300 nm på en serpentin måde på en lidt kendt proteinmatrix..
Disse sløjfer på en proteinmatrix danner en mere kompakt kromatinfiber med en diameter på 250 nm. Endelig tilpasser de sig som en enkelt spiral på 700 nm tyk, hvilket giver anledning til en af søsterkromatiderne i et mitotisk kromosom..
I sidste ende komprimerer DNA'et i kernekromatin ca. 10.000 gange på kromosomet i den delende celle. I mellemfasekernen er dens komprimering også høj, da den er ca. 1000 gange sammenlignet med det "lineære" DNA..

I en verden af udviklingsbiologi siges gametogenese at nulstille epigenomet. Det vil sige, det sletter DNA-mærkerne, at livet for den person, der gav anledning til den gamete, der blev produceret eller oplevet.
Disse tags inkluderer DNA-methylering og kovalente ændringer af histoner (kode for histoner). Men ikke hele epigenomet nulstilles. Hvad der er tilbage med mærker, vil være ansvarlig for faderens eller moderens genetiske aftryk.
Den implicitte nulstilling til gametogenese er lettere at se i sædceller. I sæd er DNA ikke pakket med histoner. Derfor nedarves informationen generelt forbundet med dens ændringer i producentorganismen ikke..
I sæd er DNA pakket gennem interaktion med ikke-specifikke DNA-bindende proteiner kaldet protaminer. Disse proteiner danner disulfidbindinger med hinanden og hjælper således med at danne overlappende DNA-lag, der ikke frastøder elektrostatisk..



Endnu ingen kommentarer