EN fylogenetisk træ det er en grafisk matematisk repræsentation af historien og forfædrenes efterkommers forhold mellem grupper, populationer, arter eller enhver anden taksonomisk kategori. Teoretisk kan alle fylogenetiske træer grupperes i livets træ og udgør det universelle træ.
Disse grafiske repræsentationer har revolutioneret studiet af evolutionærbiologi, da de giver mulighed for at etablere og definere en art, teste forskellige evolutionære hypoteser (såsom endosymbiotisk teori), evaluere sygdommens oprindelse (såsom HIV) osv..
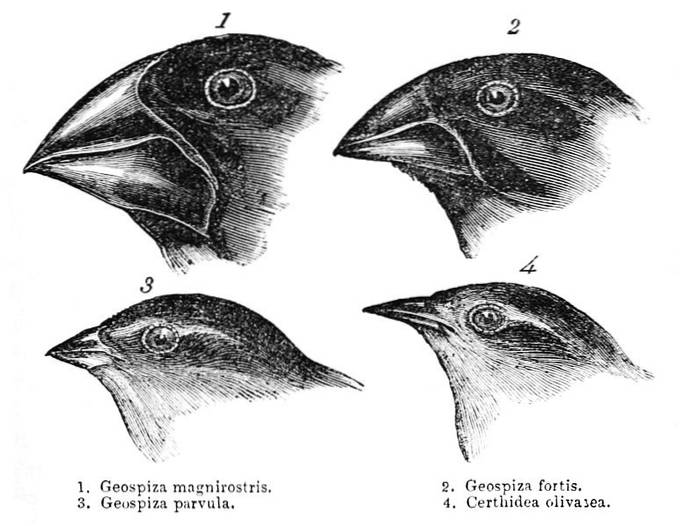
Træer kan rekonstrueres ved hjælp af morfologiske eller molekylære tegn eller begge dele. Tilsvarende er der forskellige metoder til at opbygge dem, den mest almindelige er den kladistiske metode. Dette søger at identificere delte afledte tegn, kendt som synapomorphy.
Artikelindeks
Et af de principper, der er udviklet af Charles Darwin, udgør den fælles forfædre for alle levende organismer - det vil sige at vi alle deler en fjern forfader.
På "Arternes oprindelse" Darwin rejser metaforen for "livets træ". Faktisk bruger han et hypotetisk grafisk træ til at udvikle sin idé (mærkeligt nok er det den eneste illustration af Kilde).
Repræsentationen af denne metafor er det, vi kender som fylogenetiske træer, som giver os mulighed for grafisk at vise historien og forholdet mellem en bestemt gruppe af organismer..
I fylogenetiske træer kan vi skelne mellem følgende dele - fortsætter med den botaniske analogi:
Grene: Linjerne på træet kaldes "grene", og disse repræsenterer undersøgelsespopulationerne over tid. Afhængig af typen af træ (se nedenfor) kan grenens længde måske eller ikke have en betydning.
På spidsen af grenene finder vi de organismer, som vi vil evaluere. Disse kan være enheder, der i øjeblikket er i live, eller uddøde væsener. Arten ville være bladene på vores træ.
Rod: roden er den ældste gren af træet. Nogle har det og kaldes rodfæstede træer, mens andre ikke har det.
Knuder: grenens forgreningspunkter i to eller flere slægter kaldes noder. Prikken repræsenterer den seneste fælles forfader til efterkommergrupperne (bemærk at disse forfædre er hypotetiske).
Eksistensen af en knude indebærer en specieringsbegivenhed - oprettelse af nye arter. Efter dette følger hver art sit evolutionære forløb.
Ud over disse tre grundlæggende begreber er der andre nødvendige udtryk, når det kommer til fylogenetiske træer:
Politomi: når et fylogenetisk træ har mere end to grene i en knude, siges det at der er en polytomi. I disse tilfælde løses det fylogenetiske træ ikke fuldt ud, fordi forholdet mellem de involverede organismer ikke er klart. Dette skyldes normalt mangel på data og kan kun løses, når en forsker akkumulerer mere.
Ekstern gruppe: i fylogenetiske emner er det almindeligt at høre begrebet ekstern gruppe - også kaldet udgruppe. Denne gruppe er valgt for at kunne rodfæste træet. Det skal vælges som en taxon, der tidligere afveg fra studiegruppen. For eksempel, hvis jeg studerer pighuder, kan du bruge udgruppe hav sprøjter.
Der er tre basistyper af træer: cladogrammer, additivtræer og ultrametriske træer..
Kladogrammer er de enkleste træer og viser organismernes forhold i form af fælles herkomst. Oplysningerne om denne type træ findes i forgreningsmønstrene, da størrelsen på grenene ikke har nogen yderligere betydning.
Den anden type træ er additivet, også kaldet metriske træer eller fylogrammer. Grenernes længde er relateret til mængden af evolutionær ændring.
Endelig har vi ultrametriske træer eller dendogrammer, hvor alle spidserne af træerne er i samme afstand (hvilket ikke er tilfældet i fylogrammet, hvor et tip kan se lavere eller højere ud end dets partner). Grenens længde er relateret til evolutionstiden.
Valget af træ er direkte relateret til det evolutionære spørgsmål, som vi vil besvare. For eksempel, hvis vi kun bekymrer os om forholdet mellem individer, vil et kladogram være tilstrækkeligt til undersøgelsen.
Selvom fylogenetiske træer ofte er meget anvendte grafer i evolutionær biologi (og generel biologi), er der mange studerende og praktikere, der fejlagtigt fortolker budskabet, som disse tilsyneladende enkle grafer er beregnet til at levere til læseren..
Den første fejl er at læse dem sidelæns, forudsat at evolution indebærer fremskridt. Hvis vi forstår den evolutionære proces korrekt, er der ingen grund til at tro, at de forfædres arter er til venstre og de mere avancerede arter til højre..
Selvom den botaniske analogi af et træ er meget nyttig, kommer der et punkt, hvor det ikke længere er så nøjagtigt. Der er en vigtig træstruktur, der ikke findes i træet: stammen. I fylogenetiske træer finder vi ingen hovedgrene.
Specifikt kan nogle mennesker betragte mennesket som det ultimative "mål" for evolutionen, og derfor arten. Homo sapiens skal altid være placeret som endelig enhed.
Denne opfattelse er imidlertid ikke i overensstemmelse med evolutionære principper. Hvis vi forstår, at fylogenetiske træer er mobile elementer, kan vi placere Homo i enhver terminal position af træet, da denne egenskab ikke er relevant i repræsentationen.
En vital egenskab, som vi skal forstå om fylogenetiske træer, er, at de repræsenterer ikke-statiske grafer..
I dem kan alle disse grene rotere - på samme måde som en mobil kan gøre det. Vi mener ikke, at vi kan flytte grenene efter ønske, fordi nogle bevægelser ville antyde ændring af mønsteret eller topologi Fra træet. Hvad vi kan rotere er knudepunkterne.
For at fortolke et træs budskab må vi ikke fokusere på grenene, vi skal fokusere på grenpunkterne, som er det vigtigste aspekt af grafen.
Derudover skal vi huske på, at der er flere måder at tegne et træ på. Mange gange afhænger det af stilen på bogen eller magasinet, og ændringer i filialernes form og placering påvirker ikke den information, de vil formidle til os..
Hvornår skal vi henvise til arter nuværende vi skal ikke anvende forfædres konnotationer til dem. For eksempel, når vi tænker på forholdet mellem chimpanser og mennesker, kunne vi fejlagtigt forstå, at chimpanser er forfædre med hensyn til vores slægt..
Imidlertid var den fælles forfader til chimpanser og mennesker hverken. At tro, at chimpansen er forfædres, ville være at antage, at dens udvikling stoppede, når begge slægter var adskilt..
Efter den samme logik af disse ideer fortæller et fylogenetisk træ os heller ikke, om der er unge arter. Da de allelfrekvenser er i konstant forandring, og der er nye tegn, der ændrer sig over tid, er det vanskeligt at bestemme en arts alder, og bestemt giver et træ os ikke sådanne data.
"Ændringen i allelfrekvenser over tid" er, hvordan populationsgenetik definerer evolution.
Når vi ser på et fylogenetisk træ, skal vi forstå, at denne graf simpelthen er en hypotese genereret ud fra konkrete beviser. Det kan være, at hvis vi tilføjer flere tegn til træet, ændrer det dets topologi.
Forskernes ekspertise i at vælge de bedste tegn til at belyse de pågældende organismers forhold er nøglen. Derudover er der meget kraftfulde statistiske værktøjer, der giver forskere mulighed for at evaluere træer og vælge den mest sandsynlige..
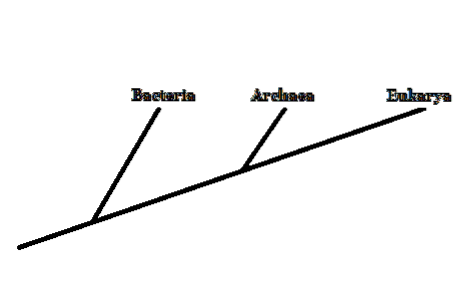
I 1977 foreslog forskeren Carl Woese at gruppere levende organismer i tre domæner: Archaea, Bacteria og Eukarya. Dette nye klassificeringssystem (tidligere var der kun to kategorier, Eukaryota og Prokaryota) var baseret på den ribosomale RNA-molekylære markør.
Bakterier og eukaryoter er almindeligt kendte organismer. Archaea forveksles ofte med bakterier. Disse adskiller sig imidlertid dybt i strukturen af deres cellulære komponenter..
Derfor, selv om de er mikroskopiske organismer som bakterier, er medlemmer af Archaea-domænet tættere beslægtet med eukaryoter - fordi de deler en tættere fælles forfader..
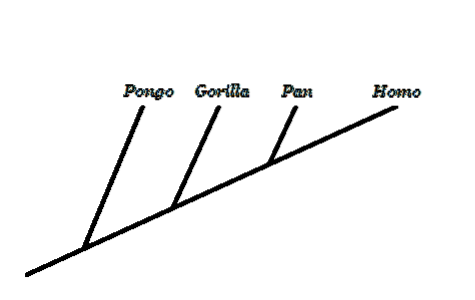
Inden for evolutionær biologi er et af de mest kontroversielle emner menneskets udvikling. For modstanderne af denne teori er en udvikling, der starter fra en apelignende forfader, der gav anledning til det moderne menneske, ikke logisk..
Et nøglebegreb er at forstå, at vi ikke udviklede os fra nuværende aber, men snarere deler vi en fælles forfader med dem. I abernes og menneskets træ skiller det sig ud, at det, vi kender som "abe", ikke er en gyldig monofyletisk gruppe, da det udelukker mennesker.
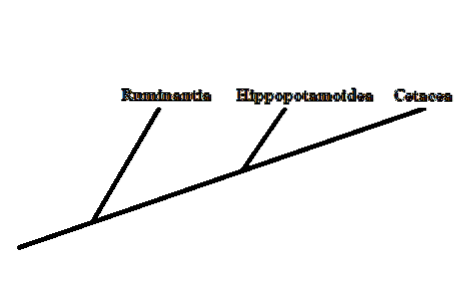
Evolutionært set repræsenterede hvaler en gruppe hvirveldyr, hvis forhold til resten af deres medpattedyr ikke var særlig klare. Morfologisk har hvaler, delfiner og andre medlemmer få ligheder med resten af pattedyrene.
I øjeblikket har det takket være studiet af forskellige morfologiske og molekylære tegn været muligt at konkludere, at søstergruppen til de store hvaler er dannet af artiodactylerne - hovdyr med jævne hov..


Endnu ingen kommentarer