Mitokondrie-DNA er et lille cirkulært DNA-molekyle, der findes inde i disse organeller i eukaryote celler. Dette lille genom koder for et meget begrænset antal proteiner og aminosyrer i mitokondrierne. Det er almindeligt at finde navnet "mitokondrie-DNA" forkortet i mange lærebøger og videnskabelige artikler som "MtDNA"eller på engelsk"mtDNA".
Mitokondrier er essentielle organeller for eukaryote celler, da de er ansvarlige for at omdanne energien fra mad, der forbruges i form af sukker til en form for energi, som celler kan bruge (for eksempel ATP).
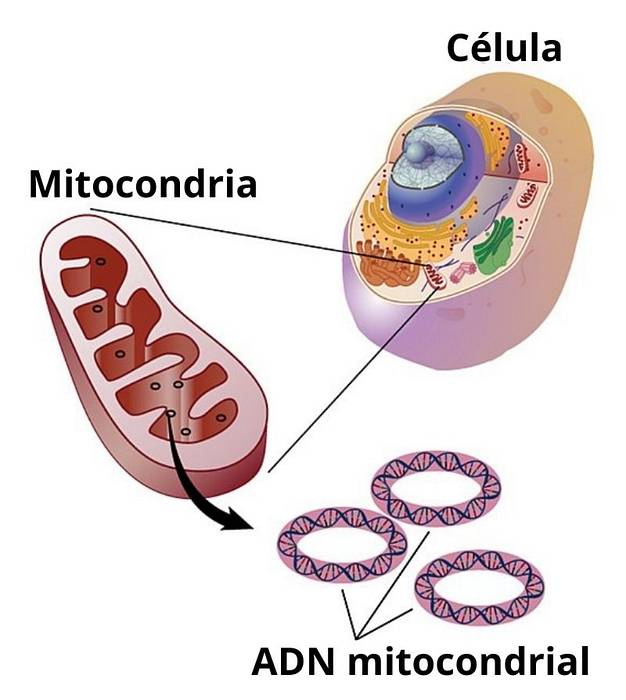
Alle celler i eukaryote organismer har mindst en mitokondrion inde i sig. Der er dog celler såsom hjertemuskelceller og skeletmuskelceller, der kan have hundreder af mitokondrier inde..
Mitokondrier har deres eget proteinsynteseapparat uafhængigt af celleapparatet med ribosomer, transfer-RNA'er og en aminoacyl-RNA-transferase-synthetase fra det indre af organellen; skønt ribosomalt RNA er mindre end cellen, der huser dem.
Et sådant apparat viser stor lighed med apparatet til proteinsyntese af bakterier. Desuden er dette apparat, som i prokaryoter, ekstremt følsomt over for antibiotika, men meget forskelligt fra proteinsyntese i eukaryote celler..
Udtrykket "mitokondrier" blev introduceret af Benda i slutningen af det 12. århundrede, og teorien om "endosymbiose" er den mest accepterede om dens oprindelse. Dette blev offentliggjort i 1967 af magasinet Lynn Margulis Tidsskrift for teoretisk biologi.
Teorien om "endosymbiose" placerer mitokondriernes oprindelse for millioner af år siden. Det teoretiseres, at en cellulær forfader til eukaryote celler "slugte" og inkorporerede en bakterielignende organisme i dens stofskifte, som senere blev det, vi nu kender som mitokondrier.
Hos pattedyr er generelt hele genomet, der omfatter mitokondrie-DNA, organiseret i et cirkulært kromosom på 15.000 til 16.000 par nukleotider eller, hvad der er det samme, på 15 til 16 Kb (kilobaser).
Inde i de fleste mitokondrier kan du få flere kopier af mitokondrie-kromosomet. I humane somatiske celler (ikke-kønsceller) er det almindeligt at finde mindst 100 kopier af mitokondrie-kromosomet.
I højere planter (angiospermer) er mitokondrie-DNA normalt meget større, for eksempel i majsplanten kan det cirkulære kromosom i mitokondrie-DNA måle op til 570 Kb.
Mitokondrie-DNA optager ca. 1% af det samlede DNA for de somatiske celler hos de fleste hvirveldyr. Det er et meget konserveret DNA i dyreriget, i modsætning til hvad der observeres i planter, hvor der er en bred mangfoldighed.
I nogle "kæmpe" eukaryote celler, såsom ovules (kvindelige kønsceller) hos pattedyr eller i celler, der indeholder mange mitokondrier, kan mitokondrie-DNA udgøre op til 1/3 af det samlede cellulære DNA.
Mitokondrie-DNA har nogle forskellige egenskaber end nukleart DNA: det har en anden tæthed og et forhold mellem guanin-cytosin (GC) og adenin-thymin (AT) basepar.
GC-basepardensiteten i mitokondrie-DNA er 1,68 g / cm3, og indholdet er 21%; mens i nukleart DNA er densiteten 1,68 g / cm3, og indholdet er omkring 40%.
Mitokondrie-DNA har mindst 37 gener, der er essentielle for normal mitokondriefunktion. Af disse 37 har 13 informationen til at producere de enzymer, der er involveret i oxidativ phosphorylering.
Disse 13 gener koder for 13 polypeptidkomponenter i enzymkomplekserne, der hører til elektrontransportkæden og er placeret i mitokondriens indre membran..
På trods af de 13 polypeptider, som mitokondrie-DNA bidrager til elektrontransportkæden, består det af mere end 100 forskellige polypeptider. Disse 13 komponenter er imidlertid vigtige for oxidativ fosforylering og elektrontransportkæden..
Blandt de 13 polypeptider, der syntetiseres fra mitokondrie-DNA, skiller I, II og III-underenhederne af cytochrom C-oxidasekomplekset sig ud og VI-underenheden af ATPase-pumper indlejret i organellens indre membran..
Oplysningerne, der er nødvendige for syntesen af resten af de komponenter, der udgør mitokondrierne, kodes af nukleare gener. Disse syntetiseres i cytoplasmaet som resten af de cellulære proteiner og importeres derefter til mitokondrier takket være specifikke signaler..
I oxidativ fosforylering anvendes ilt og sukkeratomer såsom glucose til syntese eller dannelse af adenosintriphosphat (ATP), som er den kemiske art, der anvendes af alle celler som en energikilde..
De resterende mitokondrielle gener har instruktionerne til at syntetisere transfer-RNA'er (tRNA'er), ribosomale RNA'er og enzymet aminoacyl-RNA-transferase-synthetase (tRNA), der er nødvendigt for syntesen af proteiner i mitokondrierne..
Indtil for nylig blev det antaget, at mitokondrie-DNA blev overført udelukkende gennem moderens arv, det vil sige ved direkte afstamning fra moderen..
En artikel offentliggjort af Shiyu Luo og samarbejdspartnere i bladet Forhandlinger fra National Academy of Sciences i Amerikas Forenede Stater (PNAS) i januar 2019, fandt, at mitokondrie-DNA i sjældne tilfælde kan arves fra begge forældre, både fra faren og moderen.
Før offentliggørelsen af denne artikel var det en kendsgerning for forskere, at Y-kromosomet og mitokondrie-DNA blev arvet intakt fra henholdsvis far og mor til afkom..
Den "intakte" arv af generne af Y-kromosomet i de mitokondrielle gener indebærer, at det genetiske materiale ikke gennemgår ændringer ved rekombination, og gennem årene varierer de kun på grund af spontane mutationer, så variationen er ret lav.
På grund af dette udføres de fleste populationsmobiliseringsundersøgelser på basis af disse gener, da det f.eks. Er let for genetikere at konstruere familietræer ved hjælp af mitokondrie-DNA..
Meget af menneskets historie er blevet rekonstrueret gennem den genetiske historie med mitokondrie-DNA. Mange forretningshuse tilbyder endda at afklare familiebåndet for hver levende person med deres forfædre gennem teknikker, der studerer disse egenskaber..
Den første model af mitokondrie-DNA-replikation blev foreslået i 1972 af Vinograd og samarbejdspartnere, og denne model er stadig gyldig med nogle ændringer. Generelt er modellen baseret på en envejsreplikation, der begynder med to forskellige replikationsoprindelse..
Forskere klassificerer mitokondrie-kromosomet i to forskellige kæder, den tunge kæde, H eller OH, fra engelsk "tung"Og let kæde, L eller OL fra engelsk"Lys”. Disse er identificeret og placeret i de to ikke tildelte åbne læserammer (URF) på mitokondrie-kromosomet.
Replikationen af mitokondriegenomet begynder i den tunge kæde (OH) og fortsætter i en enkelt retning, indtil den fulde længde af den lette kæde (OL) er produceret. Efterfølgende er proteiner kaldet "mitokondrie enkeltstrengede DNA-bindende proteiner" knyttet til at beskytte kæden, der fungerer som en "forælder" eller "skabelon.".
Enzymerne, der er ansvarlige for, at separationen for replikation finder sted (replikosom) passerer ind i lysbåndet (OL), og der dannes en loopstruktur, der blokerer bindingen af mitokondrie enkeltstrengede DNA-bindende proteiner.
I denne sløjfe binder mitokondrie-RNA-polymerase, og syntesen af den nye primer begynder. Overgangen til syntesen af den tunge kæde (OH) sker 25 nukleotider senere.
Lige på tidspunktet for overgangen til den tunge kæde (OH) erstattes mitokondrie-RNA-polymerase med mitokondrie-replikativ DNA-polymerase i 3'-enden, hvor replikation oprindeligt begyndte.
Endelig fortsætter syntesen af begge kæder, både tunge (OH) og lette (OL) kontinuerligt, indtil der dannes to komplette cirkulære molekyler af dobbeltstrenget DNA (dobbeltstrenget)..
Der er mange sygdomme relateret til en funktionsfejl i mitokondrie-DNA'et. De fleste er forårsaget af mutationer, der beskadiger sekvensen eller informationen i genomet.
En af de bedst studerede sygdomme, der har været direkte relateret til ændringer i mitokondrie-DNA-genomet, er høretab på grund af stigende alder.
Denne tilstand er et produkt af genetiske, miljømæssige og livsstilsfaktorer. Når mennesker bliver ældre, akkumulerer mitokondrie-DNA skadelige mutationer, såsom sletninger, translokationer, inversioner og mere.
Skader på mitokondrie-DNA skyldes hovedsageligt akkumulering af reaktive iltarter, disse er biprodukter fra energiproduktion i mitokondrier.
Mitokondrie-DNA er især sårbart over for skader, da det ikke har et reparationssystem. Derfor skader ændringer forårsaget af reaktive iltarter mitokondrie-DNA og får organellen til at fungere forkert og forårsager celledød..
Cellerne i det indre øre har et stort behov for energi. Dette krav gør dem særligt følsomme over for mitokondrie-DNA-skader. Disse skader kan ændre det indre øres funktion irreversibelt og føre til totalt høretab..
Mitokondrie-DNA er især følsomt over for somatiske mutationer, mutationer, der ikke arves fra forældre. Disse typer af mutationer forekommer i DNA fra nogle celler gennem folks liv..
Der er beviser for, at mitokondrie-DNA-ændringer som følge af somatiske mutationer forbinder med visse typer kræft, tumorer i brystkirtler, tyktarm, mave, lever og nyre..
Mutationer i mitokondrie-DNA har også været forbundet med blodkræft såsom leukæmi og lymfomer (kræft i immunsystemets celler).
Specialister forbinder somatiske mutationer i mitokondrie-DNA med en stigning i produktionen af reaktive iltarter, faktorer der øger mitokondrie-DNA-skader og skaber manglende kontrol i cellevækst.
Man ved ikke meget om, hvordan disse mutationer øger den ukontrollerede celledeling af celler, og hvordan de ender med at udvikle sig som kræfttumorer..
Nogle tilfælde af cyklisk opkastning, der er typisk for barndommen, menes at være relateret til mutationer i mitokondrie-DNA. Disse mutationer forårsager tilbagevendende episoder med kvalme, opkastning og træthed eller sløvhed..
Forskere forbinder disse opkastningsepisoder med det faktum, at mitokondrier med beskadiget mitokondrie-DNA kan påvirke visse celler i det autonome nervesystem, hvilket påvirker funktioner som puls, blodtryk og fordøjelse.
På trods af disse foreninger er det endnu ikke klart, hvordan ændringer i mitokondrie-DNA forårsager tilbagevendende episoder af cyklisk opkastningssyndrom..


Endnu ingen kommentarer