
Det ionkanaler de er hule membranstrukturer, der danner ledninger eller porer, der krydser tykkelsen af membranen og kommunikerer det ydre af cellen med dens cytosol og omvendt; nogle kan have et portsystem, der regulerer åbningen.
Disse kanaler er fyldt med vand og styrer passagen af specifikke ioner fra den ene side af membranen til den anden. De er dannet af nogle proteiner, der er typiske for cellemembraner, der danner cylindriske rørformede strukturer, der krydser dem i bredden..
Transportmekanismerne over disse membraner kan bredt klassificeres i passiv eller aktiv transport. Passive er dem, der tillader passage af stoffer til fordel for deres koncentrationsgradienter, mens aktive kræver energiforbrug, da de bevæger stoffer mod deres koncentrationsgradienter..
Ionkanaler udgør en passiv transportmekanisme, der kan klassificeres efter deres specificitet, det vil sige efter den type ion, som de tillader at passere igennem, eller efter den måde, hvorpå de åbnes eller lukkes..

Hovedfunktionen for disse membrantransportsystemer er at tillade reguleret passage af stoffer ind i eller ud af celler og således opretholde intracellulære koncentrationer af ioner og andre stoffer..
Tilstedeværelsen af cellemembraner og ionkanaler er afgørende for opretholdelsen af koncentrationsforskelle mellem det intracellulære og det ekstracellulære medium, hvilket er relevant fra mange synsvinkler..
Ionkanaler, især de afhængige ligander, er meget vigtige inden for farmakologi og medicin, da mange medikamenter kan efterligne funktionerne af naturlige ligander og således binder til kanalen, åbner eller lukker den, alt efter tilfældet..
Andre lægemidler er i stand til at blokere bindingsstedet og dermed forhindre virkningen af den naturlige ligand.
Artikelindeks
Strukturen af ionkanaler består af specifikke transmembranproteiner, der har en rørform og efterlader en pore eller et hul, der muliggør kommunikation mellem indersiden og ydersiden af cellen eller mellem intracellulære rum (organeller)..
Hver ionkanal involverer et specifikt strukturelt membranprotein, og der er beskrevet mere end 100 gener, der koder for specifikke ionkanaler..
For natriumkanalen kaldes for eksempel 10 gener SCN der koder for forskellige proteiner fordelt i forskellige væv med særlige funktioner og strukturer.
Ligeledes er der beskrevet en betydelig mængde gener, der koder for forskellige proteiner, der udgør kaliumkanaler, der hører til forskellige familier og har forskellige aktiverings-, åbnings- og inaktiveringsmekanismer..
Typisk er en funktionel ionkanal forbundet med en membran sammensat af samlingen af 4 til 6 lignende polypeptidsubenheder (homo-oligomerer) eller forskellige (hetero-oligomerer), der danner en central pore imellem dem..
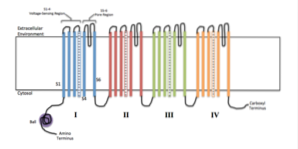
Hver underenhed varierer alt efter kanalens karakteristika og egenskaber, da mange er specifikke for visse ioner og har forskellige åbnings- og lukningsmekanismer..
Nogle kanaler er sammensat af en enkelt polypeptidkæde organiseret i gentagne motiver, der går igennem flere gange tykkelsen af membranen og fungerer som ækvivalent med en proteinunderenhed..
Ud over disse underenheder, der i litteraturen er kendt som α-underenheder, har nogle ionkanaler også en eller flere hjælpeenheder (ß eller γ), der regulerer deres åbning og lukning..
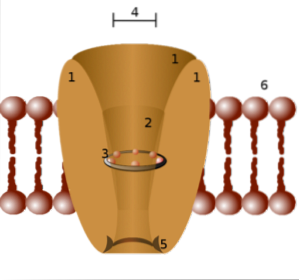
Specificiteten af hver kanal er relateret til diameteren på den pore, der dannes af transmembranproteinerne og sidekæderne (─R) af de aminosyrer, der udgør dem..
På denne måde er der kanaler, der kun lader natrium, kalium, calciumioner passere osv., Da sidekæderne fungerer som en "sigte".
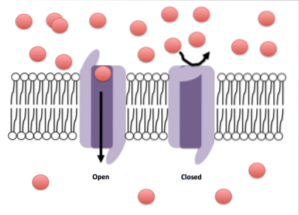
Et andet vigtigt træk ved mange kanaler er portene. Kanaler med disse egenskaber kan åbnes eller lukkes mod lokale ændringer, der opstår i membranmikromiljøet, der omgiver kanalen..
Afhængigt af kanaltypen kan disse ændringer være mekaniske, termiske (temperaturændringer), elektriske (spændingsændringer) eller kemiske (binding af en ligand).
I de såkaldte passive ionkanaler, som er dem, der forbliver åbne og tillader specifik passage af visse ioner, har disse strukturer ikke porte eller er følsomme over for ligander eller andre typer stimuli..
I andre ionkanaler, som er følsomme over for tilstedeværelse eller binding af ligander, er der et bindingssted for ligand enten på den ekstracellulære side eller mod cellecytosolen, og i disse tilfælde har porerne eller kanalerne en gate, der kan åbnes eller lukkes alt efter status for din ligand.
I tilfælde af at have et ligandsted i den intracellulære del har disse kanaler normalt anden budbringere som ligander. Et eksempel på ionkanaler, der åbnes eller lukkes af anden messenger-mekanisme, er olfaktoriske receptorer:
Duftmolekyler binder til deres receptorer på den ekstracellulære side. Disse receptorer er til gengæld bundet til et aktiveret G-protein, der igen aktiverer protein-adenylcyclasen, der danner cAMP, som er en anden messenger..
CAMP'en binder til et intracellulært bindingssted for nogle calciumkanaler, hvilket resulterer i dets åbning og indtrængen af calcium i cellen.
Som om det var en dominoeffekt, binder calcium sig til et ligandsted i en anden klorkanal, som genererer dets åbning og udgang af denne ion og forårsager depolarisering af olfaktoriecellen.
Det er vigtigt at fremhæve, at de ændringer, der genereres af liganderne eller stimuli, der påvirker ionkanalerne, svarer til konformationsændringer af proteinerne, der udgør strukturen i kanalen..
Med andre ord er de konformationsændringer, der kan bevæge en port og lukke eller åbne en kanal, intet andet end at nærme sig eller distancere sig for de proteinunderenheder, der komponerer den..
Nogle kanaler, især spændingsafhængige kanaler, kan komme ind i en ildfast tilstand, hvor den samme spændingsændring, som aktiverede dem, nu ikke længere aktiverer dem..
For eksempel i spændingsstyrede calciumkanaler åbner spændingsændringen kanalen, og calcium kommer ind, og når den er inde i cellen, binder den samme ion til et calciumkanalbindingssted, og calciumkanalen lukkes..
En anden form for reversibel inaktivering af calciumkanalen, der forklarer dets ildfasthed efter aktivering, er kanalphosphorylering på grund af øget intern calciumkoncentration..
Det vil sige en calciumkanal kan inverteres irreversibelt på grund af tilstedeværelsen af patologisk høje koncentrationer af ionen, som medierer rekrutteringen af spaltningsenzymer af andre calciumaktiverede proteiner..
Ligandregulerede kanaler kan komme ind i en ildfast tilstand, når de udsættes for deres ligand i lang tid, idet denne mekanisme kaldes desensibilisering..
Narkotika, gift og toksiner kan påvirke reguleringen af ionkanaler, lukke eller holde dem åbne eller i nogle tilfælde optage stedet for liganden og dermed forstyrre dens funktion..
Ionkanaler har en række funktioner, direkte eller indirekte.
- De er ansvarlige for at regulere strømmen af ioner gennem plasma og organellære membraner i alle celler..
- De tillader eksistensen af en kontrol over de forskellige ioners intracellulære koncentrationer.
- I neuroner og muskelceller styrer ionkanaler variationerne i membranpotentiale, der opstår under handlingspotentialer og under postsynaptiske effektorceller..
- De calciumkanaler, der genererer nettoflow af calcium i det intracellulære rum, er ansvarlige for aktiveringen af adskillige enzymer og proteiner, der deltager i mange metaboliske processer.
- Ligeledes initierer stigningen i calcium på grund af en stigning i dets transport frigivelsesmekanismen for neurotransmittere til det synaptiske rum af neuroner..
- Derfor er ionkanalernes funktion også relateret til mekanismerne til cellulær kommunikation..
Som nævnt ovenfor kan membrantransportmekanismer være aktive eller passive afhængigt af, om de bruger energi fra cellen, hvor de er placeret eller ej. Passive mekanismer klassificeres i simpel diffusion og letter diffusion.
Enkel diffusion tillader passage gennem fosfolipidstrukturen i membranen af fedtopløselige molekyler af lille størrelse med apolære egenskaber og uden opladning..
Således passerer f.eks. Gasser såsom ilt (O2) og kuldioxid (CO2), ethanol og urinstof, for at nævne nogle få, til fordel for deres koncentrationsgradient.
Let diffusion er en, der lettes af proteiner, og der er to typer af denne passive transportmekanisme: ionkanaler og transportproteiner eller transportproteiner..
Ionkanaler er den mekanisme, der bruges mest af celler til transport af ioner, der ikke kan passere ved simpel diffusion, enten fordi de har en elektrisk ladning, og fosfolipiderne i membranen afviser dem på grund af deres størrelse og polaritet eller andre egenskaber..
Diffusion muliggjort af transportørproteiner bruges til at transportere større stoffer med eller uden opladning, såsom glukose og andet sukker..
Aktiv membrantransport er den, der forekommer mod koncentrationsgradienten af det opløste stof, der transporteres og kræver energiforbrug i form af ATP. Blandt transportører af denne type er pumper og vesikulær transport.
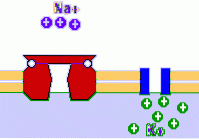
Som et eksempel på pumperne er natrium / kaliumpumpen, som fjerner tre natrium og indfører to kalium. Der er også calciumpumper.
Eksempler på vesikulær transport er endocytose, exocytose, pinocytose og fagocytose; alle disse aktive transportmekanismer.
Fra dette punkt henvises der til ionkanaler, der tillader passage af ioner gennem en membran til fordel for deres koncentrationsgradienter, dvs. de er passive transportkanaler..
Generelt er hver af disse kanaler specifik for en enkelt ion med undtagelse af et par kanaler, der tillader transport af par af ioner..
En måde at klassificere ionkanaler på er ved at gruppere dem efter den mekanisme, der er ansvarlig for deres åbning. Således er passive kanaler, spændingsregulerede kanaler (spændingsafhængige), ligandregulerede kanaler og kanaler reguleret af mekaniske stimuli blevet beskrevet..
- Passive kanaler: de er kanaler, der er permanent åbne og ikke reagerer på nogen form for stimulus; disse er specifikke for visse ioner.
- Spændingsafhængige kanalerDisse kan åbne eller lukke (afhængigt af kanalen) i lyset af ændringer i membranspændingen. De er meget vigtige for cellesignalering, især i pattedyrets centralnervesystem.
- Ligandafhængige kanaler: Også kaldet ligand-gated eller ligand-regulerede kanaler, de er bredt fordelt i forskellige menneskelige kropsceller, men i nervesystemet udgør de de ionkanaler aktiveret af neurotransmittere og er essentielle for synaptisk transmission og intercellulær signalering.
Eksempel på ligandafhængige ionkanaler aktiveret af neurotransmittere er natrium / kaliumkanaler aktiveret af glutamat..
Aktivering af kolinerge receptorer, i dette tilfælde binding af acetylcholin til den postsynaptiske membran (kanalligand), åbner ligandafhængige natriumkanaler og tillader indgang af denne ion efter dens koncentrationsgradient..
- Kanaler reguleret af mekaniske stimuli: Dette er kanaler, der kan aktiveres ved belastning eller tryk. Disse mekaniske kræfter overføres til kanalen gennem cytoskelettet, og kanalen åbnes.


Endnu ingen kommentarer