Det cytoskelet det er en cellulær struktur sammensat af filamenter. Det er spredt gennem cytoplasmaet, og dets funktion er hovedsageligt understøttende for at opretholde arkitekturen og celleformen. Strukturelt er den sammensat af tre typer fibre, klassificeret efter deres størrelse.
Disse er actinfibre, mellemfilamenter og mikrotubuli. Hver enkelt tildeler netværket en bestemt ejendom. Celleindretningen er et miljø, hvor forskydning og transit af materialer sker. Cytoskeletet medierer disse intracellulære bevægelser.
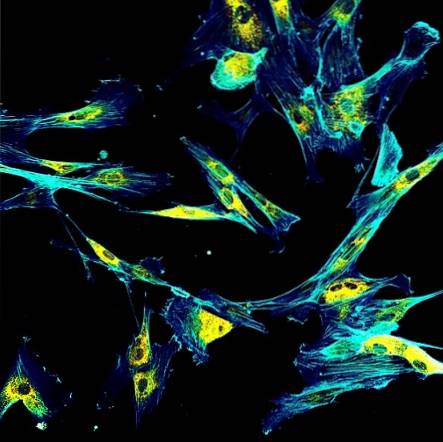
For eksempel er organeller - såsom mitokondrier eller Golgi-apparatet - statiske i det cellulære miljø; disse bevæger sig ved hjælp af cytoskeletet som en vej.
Selvom cytoskeletet klart dominerer i eukaryote organismer, er der rapporteret om en analog struktur i prokaryoter..
Artikelindeks
Cytoskeletet er en ekstremt dynamisk struktur, der repræsenterer et "molekylært stillads". De tre typer filamenter, der udgør det, er gentagne enheder, der kan danne meget forskellige strukturer afhængigt af, hvordan disse grundlæggende enheder kombineres..
Hvis vi ønsker at skabe en analogi med det menneskelige skelet, svarer cytoskelet til knoglesystemet og derudover til muskelsystemet.
Imidlertid er de ikke identiske med en knogle, da komponenterne kan samles og desintegreres, hvilket muliggør ændringer i form og giver cellen plasticitet. Cytoskeletale komponenter er ikke opløselige i vaskemidler.
Som navnet antyder, er cytoskeletets "intuitive" funktion at give cellen stabilitet og form. Når filamenterne kombineres i dette indviklede netværk, giver det cellen egenskaben til at modstå deformation.
Uden denne struktur ville cellen ikke være i stand til at opretholde en bestemt form. Det er imidlertid en dynamisk struktur (i modsætning til det menneskelige skelet), der giver cellerne egenskaben til at ændre form.
Mange af de cellulære komponenter er knyttet til dette netværk af fibre spredt i cytoplasmaet, hvilket bidrager til deres rumlige arrangement..
En celle ligner ikke en suppe med forskellige elementer flydende drift; det er heller ikke en statisk enhed. Tværtimod er det en organiseret matrix med organellerne placeret i bestemte områder, og denne proces sker takket være cytoskeletet.
Cytoskelet er involveret i bevægelse. Dette sker takket være motoriske proteiner. Disse to elementer kombinerer og tillader bevægelser i cellen.
Det deltager også i fagocytose-processen (en proces, hvor en celle fanger en partikel fra det ydre miljø, som måske eller måske ikke er mad).
Cytoskelet giver cellen mulighed for at forbinde med sit ydre miljø, fysisk og biokemisk. Denne forbindelsesrolle er det, der tillader dannelse af væv og celleforbindelser..
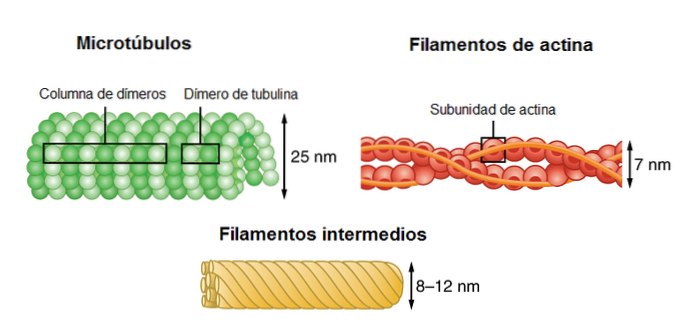
Cytoskeletet består af tre forskellige typer filamenter: actin, mellemfilamenter og mikrotubuli..
En ny kandidat foreslås i øjeblikket som en fjerde streng af cytoskelen: septin. Hver af disse dele er beskrevet detaljeret nedenfor:
Actin filamenter har en diameter på 7 nm. De er også kendt som mikrofilamenter. De monomerer, der udgør filamenterne, er ballonformede partikler.
Selvom de er lineære strukturer, har de ikke form af en "bar": de roterer på deres akse og ligner en helix. De er knyttet til en række specifikke proteiner, der regulerer deres adfærd (organisation, placering, længde). Der er mere end 150 proteiner, der er i stand til at interagere med actin.
Ekstreme kan differentieres; den ene kaldes plus (+) og den anden minus (-). I disse ender kan glødetråden vokse eller forkorte. Polymerisering er mærkbart hurtigere, jo mere ekstrem; for at der kan forekomme polymerisering, kræves ATP.
Actin kan også være som en monomer og fri i cytosolen. Disse monomerer er bundet til proteiner, der forhindrer deres polymerisering.
Actin filamenter har en rolle relateret til cellebevægelse. De tillader forskellige celletyper, både encellede og multicellulære organismer (et eksempel er immunsystemets celler), at bevæge sig i deres omgivelser.
Actin er kendt for sin rolle i muskelsammentrækning. Sammen med myosin grupperer de sig i sarkomerer. Begge strukturer muliggør en sådan ATP-afhængig bevægelse..
Den omtrentlige diameter af disse filamenter er 10 µm; deraf navnet "mellemliggende". Dens diameter er mellemliggende i forhold til de to andre komponenter i cytoskeletet.
Hver glødetråd er struktureret som følger: et ballonformet hoved ved N-terminalen og en lignende formet hale ved kulstofterminalen. Disse ender er forbundet med hinanden ved hjælp af en lineær struktur, der består af alfa-helixer..
Disse "strenge" har kugleformede hoveder, der har den egenskab, at de afvikles med andre mellemliggende filamenter, hvilket skaber tykkere sammenflettede elementer..
Mellemliggende filamenter er placeret i hele cellecytoplasmaet. De strækker sig til membranen og er ofte fastgjort til den. Disse filamenter findes også i kernen og danner en struktur kaldet "nuklear lamina".
Denne gruppe klassificeres igen i undergrupper af mellemliggende filamenter:
- Keratin filamenter.
- Vimentin filamenter.
- Neurofilamenter.
- Atomfolier.
De er ekstremt stærke og modstandsdygtige elementer. Faktisk, hvis vi sammenligner dem med de to andre filamenter (actin og mikrotubuli), får de mellemliggende filamenter stabilitet..
Takket være denne egenskab er dens hovedfunktion mekanisk, der modstår cellulære ændringer. De findes rigeligt i celletyper, der oplever konstant mekanisk stress; for eksempel i nerve-, epitel- og muskelceller.
I modsætning til de to andre komponenter i cytoskelettet kan de mellemliggende filamenter ikke samles og løsnes i deres polære ender..
De er stive strukturer (for at være i stand til at udføre deres funktion: cellulær understøtning og mekanisk reaktion på stress) og samling af filamenterne er en proces, der er afhængig af phosphorylering..
De mellemliggende filamenter danner strukturer kaldet desmosomer. Sammen med en række proteiner (cadheriner) skabes disse komplekser, der danner krydset mellem celler.
Mikrotubuli er hule elementer. De er de største filamenter, der udgør cytoskelettet. Diameteren af mikrorørene i dens indre del er omkring 25 nm. Længden er ret variabel inden for området 200 nm til 25 µm.
Disse filamenter er uundværlige i alle eukaryote celler. De kommer frem (eller er født) fra små strukturer kaldet centrosomer, og derfra strækker de sig ud til cellekanterne i modsætning til de mellemliggende filamenter, der strækker sig gennem det cellulære miljø..
Mikrotubuli består af proteiner kaldet tubuliner. Tubulin er en dimer, der består af to underenheder: α-tubulin og β-tubulin. Disse to monomerer er forbundet med ikke-kovalente bindinger.
En af dens mest relevante egenskaber er evnen til at vokse og forkorte, idet de er ret dynamiske strukturer, som i actinfilamenter.
De to ender af mikrotubuli kan skelnes fra hinanden. Af denne grund siges det, at der er en "polaritet" i disse filamenter. Ved hver af ekstremerne - kaldet plus p positiv og minus eller negativ - opstår processen med selvsamling.
Denne proces til samling og nedbrydning af filamentet giver anledning til et fænomen af "dynamisk ustabilitet".
Mikrotubuli kan danne meget forskellige strukturer. De deltager i processerne med celledeling og danner den mitotiske spindel. Denne proces hjælper hver dattercelle med et lige antal kromosomer.
De danner også de pisklignende vedhæng, der bruges til cellemobilitet, såsom cilia og flagella..
Mikrotubuli fungerer som veje eller "motorveje", hvor forskellige proteiner, der har transportfunktioner, bevæger sig. Disse proteiner er klassificeret i to familier: kinesiner og dyneiner. De kan rejse lange afstande inden i cellen. Transport over korte afstande udføres generelt på actin.
Disse proteiner er "fodgængere" på mikrotubuliveje. Dens bevægelse ligner en tur på mikrotubuli.
Transport involverer bevægelse af forskellige typer af elementer eller produkter, såsom blærer. I nerveceller er denne proces velkendt, fordi neurotransmittere frigives i vesikler..
Mikrotubuli deltager også i mobilisering af organeller. Især Golgi-apparatet og det endosplasmiske retikulum afhænger af disse filamenter for at indtage deres rette position. I fravær af mikrotubuli (i eksperimentelt muterede celler) ændrer disse organeller markant deres position.
I de foregående afsnit blev cytoskelet af eukaryoter beskrevet. Prokaryoter har også en lignende struktur og har komponenter, der er analoge med de tre fibre, der udgør det traditionelle cytoskelet. Til disse filamenter føjes en af sine egne bakterier: MinD-ParA-gruppen.
Cytoskeletets funktioner i bakterier svarer meget til de funktioner, de udfører i eukaryoter: støtte, celledeling, vedligeholdelse af celleform, blandt andre..
Klinisk har komponenter i cytoskelet været forbundet med kræft. Da de griber ind i delingsprocesserne, betragtes de som "mål" for at forstå og angribe ukontrolleret celleudvikling.



Endnu ingen kommentarer