Det kromatin Det er det kompleks, der dannes af DNA og proteiner, der er unikt i eukaryote organismer. Proportionelt indeholder den næsten dobbelt så mange proteiner som det gør genetisk materiale. De vigtigste proteiner i dette kompleks er histoner - små, positivt ladede proteiner, der binder til DNA gennem elektrostatiske interaktioner. Derudover har kromatin mere end tusind andre proteiner end histoner..
Den grundlæggende enhed af kromatin er nukleosomet, som består af foreningen af histoner og DNA. Dette arrangement minder om perlerne på en halskæde. Efter at have gennemgået alle de højere niveauer af DNA-organisation, når vi kromosomerne.
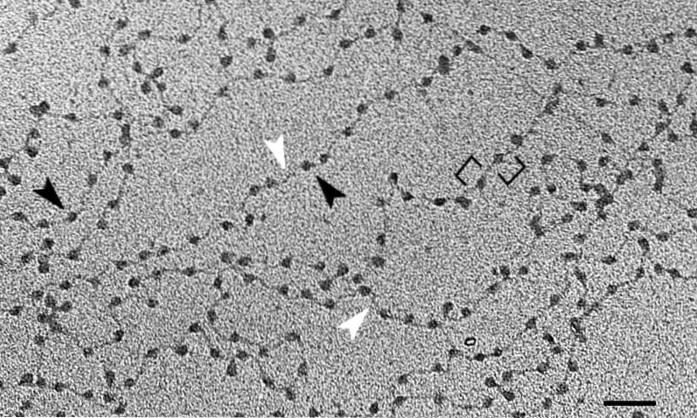
Kromatinstruktur er nært beslægtet med styringen af genekspression. Der er to hovedtyper: euchromatin og heterochromatin.
Euchromatin er kendetegnet ved at have en lav grad af komprimering, hvilket oversættes til høje niveauer af transkription. I modsætning hertil er heterochromatin transkriptionelt inaktiv på grund af dets høje grad af komprimering..
Strukturelt er der visse epigenetiske mærker af histoner, der er typiske for begge typer kromatiner. Mens euchromatin er forbundet med acetylering, er heterochromatin forbundet med et fald i kemiske grupper.
Der er nogle regioner af heterochromatin med strukturelle funktioner, såsom telomerer og centromerer.
Artikelindeks
Undersøgelsen af genetisk materiale og dets strukturelle organisation begyndte i 1831, da forskeren Robert Brown beskrev kernen. Et af de øjeblikkelige spørgsmål til denne opdagelse var at forhøre sig om denne strukturs biologiske og kemiske natur..
Disse spørgsmål begyndte at blive belyst mellem 1870 og 1900 med eksperimenterne fra Friedrich Miescher, der introducerede ordet nuclein. Walther Flemming ændrer imidlertid udtrykket og bruger kromatin at henvise til det nukleare stof.
Efterhånden som tiden går, begynder du at få en dybere forståelse af det genetiske materiale og dets egenskaber. Det var først i 1908, da den italienske forsker Pasquale Baccarini bemærkede, at kromatinet ikke var homogent og formåede at visualisere små kroppe inde i kernen..
Typerne af kromatiner - euchromatin og heterochromatin - blev oprindeligt foreslået af Emil Heitz i 1928. For at etablere denne klassificering påberåbte Heitz sig brugen af pletter.
I 1974 foreslog biologen Roger Kornberg en model til organisering af genetisk materiale i strukturer kendt som nukleosomer, en hypotese bekræftet empirisk af eksperimenterne med Markus Noll..
Chromatin er et nukleoprotein dannet ved forening af genetisk materiale - DNA - med et heterogent sæt proteiner. Denne tilknytning er meget dynamisk og erhverver en kompleks tredimensionel konformation, der gør det muligt for den at udføre sine regulatoriske og strukturelle funktioner..
Et af de vigtigste kromatinproteiner er histoner, som findes i næsten samme andel som DNA.
Histoner er grundlæggende proteiner, bemærkelsesværdigt konserverede gennem organiske væseners evolutionære historie - det vil sige, at vores histoner ikke varierer meget sammenlignet med dem fra andre pattedyr, endda et andet dyr mere fjernt fylogenetisk.
Ladningen af histoner er positiv, så de kan interagere gennem elektrostatiske kræfter med den negative ladning af phosphatrygraden til stede i DNA. Der er fem typer histoner, nemlig: H1, H2A, H2B, H3 og H4.
Der er også et antal proteiner af en anden art end histoner, der deltager i DNA-komprimering..
Den basale enhed af kromatin er nukleosomer - gentagne strukturer, der består af DNA og histoner, en konformation, der findes i hele genetisk materiale..
Den dobbelte DNA-helix er viklet ind i et kompleks af otte histoner kendt som histonoktamer. Molekylet vikles i cirka to omgange efterfulgt af et kort område (mellem 20 og 60 basepar), der adskiller nukleosomerne fra hinanden..
For at forstå denne organisation skal vi tage højde for, at DNA-molekylet er ekstremt langt (ca. 2 meter) og skal vikles på en ordnet måde for at etablere sig i kernen (hvis diameter er 3 til 10 µm). Derudover skal den gøres tilgængelig til replikering og transkription..
Dette mål opnås med forskellige niveauer af DNA-komprimering, hvor den første er de førnævnte nukleosomer. Disse ligner perlerne på en perlekæde. Ca. 150 basepar DNA er viklet derhjemme "tæller".
Der er ingen sande historier i bakterier. Derimod er der en række proteiner, der ligner histoner, og det antages, at disse bidrager til emballering af bakterielt DNA..
Organiseringen af kromatin er ikke begrænset på nukleosomniveauet. Denne tilknytning af proteiner og DNA er grupperet i en tykkere struktur på ca. 30 nm - på grund af denne tykkelse kaldes det "30 nm fiber" -niveauet..
Kromatinet organiseret i tykkelsen på 30 nm er organiseret i sin tur i form af sløjfer, der strækker sig i en slags stillads af proteinnatur (ikke histoner).
Dette er den model, der i øjeblikket anvendes, selv om der kan forventes mere komplekse komprimeringsmekanismer. Den endelige organisation består af kromosomet.
Komprimering og organisering af genetisk materiale er afgørende for flere biologiske funktioner. Forskellige medicinske tilstande er blevet forbundet med fejl i kromatinstruktur, herunder X-bundet alfa-thalassæmi, Rubinstein-Taybi syndrom, Coffin-Lowry syndrom, Rett syndrom, blandt andre..
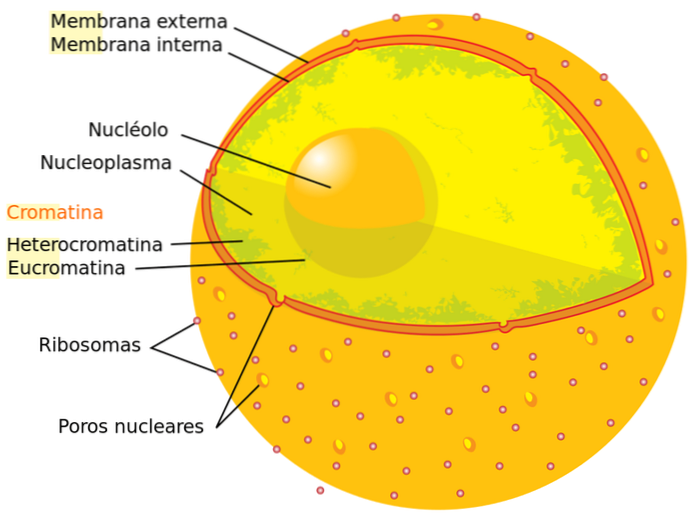
Der er to typer kromatin i cellen, afsløret ved anvendelse af pletter: euchromatin ("ægte" kromatin) og heterochromatin. I det første tilfælde observeres farvningen svagt, mens i det andet er farvningen intens.
Denne strukturelle organisation af DNA er unik for eukaryote organismer og er afgørende for kromosomernes opførsel og reguleringen af genekspression..
Hvis vi vurderer andelene af begge typer kromatiner i en celle, der er i mellemfase, finder vi, at ca. 90% af kromatinet er euchromatin, og de resterende 10% svarer til heterochromatin. Vi beskriver hver type i detaljer nedenfor:
Den væsentligste forskel, der findes mellem begge typer kromatin, er relateret til graden af komprimering eller "pakning" af molekylet i specifikke stadier i celledeling..
Selvom det genetiske materiale ved grænsefladen ser ud til at være spredt tilfældigt, er det ikke på denne måde.
Der er en betydelig organisation på dette stadium, hvor en differentiel partition af det kromosomale materiale kan ses inde i kernen.
Kromosomernes DNA fletter sig ikke sammen med andre kromosomers DNA-streng og forbliver i specifikke regioner kaldet kromosomale territorier. Denne organisation ser ud til at bidrage til genekspression.
Heterochromatin er stærkt kondenseret, hvilket gør det utilgængeligt for transkriptionsmaskineriet - så det transskriberes ikke. Derudover er det dårligt med hensyn til antallet af gener, det har..
Visse regioner af heterochromatin er vedvarende gennem cellelinier - dvs., for evigt det vil opføre sig som heterochromatin. Denne type heterochromatin er kendt som konstitutiv. Et eksempel på dette er de kondenserede regioner af kromosomer kaldet centromerer og telomerer..
Derimod er der dele af heterochromatin, der kan variere komprimeringsniveauet som reaktion på ændringer i udviklingsmønstre eller miljøvariabler..
Takket være nye undersøgelser reformeres denne vision, og nu er der tegn på, at konstituerende heterochromatin også er dynamisk og i stand til at reagere på stimuli..
En af de faktorer, der bestemmer strukturen af kromatin, er de kemiske modifikationer af histoner. I tilfælde af kromatin, der er transkriptionelt inaktivt, udviser de hypoacetylerede histoner.
Faldet i antallet af acetylgrupper er forbundet med gendæmpning, da det ikke maskerer den positive ladning af lysiner, hvilket tillader en stærk elektrostatisk interaktion mellem DNA og histoner..
Et andet epigenetisk mærke er methylering. Da tilsætningen af en methylgruppe imidlertid ikke ændrer proteinets ladning, er dens konsekvens (aktivering eller deaktivering af gener) ikke så indlysende og vil afhænge af det område af histonen, hvor mærket findes..
Empirisk har det vist sig, at methyleringen af H3K4me3 og H3K36me3 er associeret med genaktivering og den af H3K9me3 og H3K27me3..
I eksemplet med konstitutiv heterochromatin nævner vi centromeren. Denne kromosomale region har en strukturel rolle og bidrager til bevægelsen af kromosomer under både mitotiske og meiotiske celledeling begivenheder..
I modsætning til heterochromatin er euchromatin et mindre kompakt molekyle, så transkriptionsmaskineriet har let adgang (specifikt RNA-polymeraseenzymet) og kan udtrykkes aktivt via genetiske veje.
Nukleosomstrukturen i et transkriptionelt aktivt kromatin er karakteriseret ved acetylerede histoner. Tilstedeværelsen af monomethylerede lysiner er også forbundet med genaktivering.
Tilsætningen af en acetylgruppe til disse histonlysinrester neutraliserer den positive ladning af nævnte aminosyre. Den umiddelbare konsekvens af denne ændring er reduktionen af elektrostatiske interaktioner mellem histon og DNA, hvilket genererer en løsere kromatin.
Denne strukturelle modifikation tillader interaktioner mellem det genetiske materiale og transkriptionsmaskineriet, som er karakteriseret ved at være særligt omfangsrige..
Euchromatin omfatter alle gener, der er aktive og formår at nås ved hjælp af det enzymatiske maskineri relateret til transkription. Derfor er funktionerne lige så brede som de involverede gener..



Endnu ingen kommentarer