Det helikase henviser til en gruppe enzymer af den proteinhydrolytiske type, der er meget vigtige for alle levende organismer; de kaldes også motoriske proteiner. Disse bevæger sig gennem cellecytoplasmaet og omdanner kemisk energi til mekanisk arbejde gennem ATP-hydrolyse..
Dens vigtigste funktion er at bryde hydrogenbindingerne mellem de nitrogenholdige baser af nukleinsyrer og derved muliggøre deres replikering. Det er vigtigt at understrege, at helikaser er praktisk talt allestedsnærværende, da de er til stede i vira, bakterier og i eukaryote organismer..
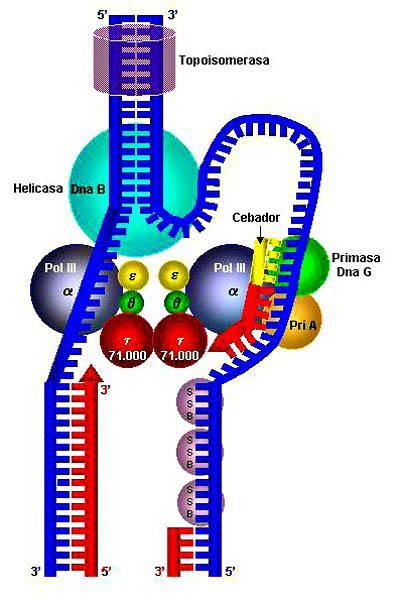
Den første af disse proteiner eller enzymer blev opdaget i 1976 i bakterier Escherichia coli; to år senere blev den første helikase opdaget i en eukaryot organisme i liljeplanter.
I øjeblikket er helikase-proteiner blevet karakteriseret i alle naturlige kongeriger inklusive vira, hvilket indebærer, at der er genereret en bred viden om disse hydrolytiske enzymer, deres funktioner i organismer og deres mekaniske rolle..
Artikelindeks
Hellicaser er biologiske eller naturlige makromolekyler, der fremskynder kemiske reaktioner (enzymer). De er hovedsageligt karakteriseret ved at adskille kemiske komplekser af adenosintriphosphat (ATP) gennem hydrolyse.
Disse enzymer bruger ATP til at binde og ombygge komplekser af deoxyribonukleinsyrer (DNA) og ribonukleinsyrer (RNA)..
Der er mindst 2 typer helikaser: DNA og RNA.
DNA-helikaser virker i DNA-replikation og er karakteriseret ved at adskille dobbeltstrenget DNA i enkelte tråde.
Disse enzymer virker i de metaboliske processer af ribonukleinsyre (RNA) og i multiplikation, reproduktion eller ribosomal biogenese.
RNA-helicase er også nøglen til præ-splejsningsprocessen af messenger RNA (mRNA) og initieringen af proteinsyntese efter transkription af DNA til RNA i cellekernen..
Disse enzymer kan differentieres i henhold til deres aminosyresekventeringshomologi med kerneaminosyren ATPase-domæne eller ved delte sekventeringsmotiver. Ifølge klassificeringen er disse grupperet i 6 superfamilier (SF 1-6):
Enzymerne i denne superfamilie har en 3'-5 'eller 5'-3' translokationspolaritet og danner ikke ringstrukturer..
Det er kendt som den største gruppe helikaser og består hovedsageligt af RNA-helikaser. De har en translokationspolaritet generelt 3'-5 'med meget få undtagelser.
De har ni motiver (fra engelsk motiver, som er oversat som "tilbagevendende elementer") af stærkt konserverede aminosyresekvenser og, ligesom SF1, ikke danner ringstrukturer.
De er karakteristiske helikaser af vira og har en unik 3'-5 'translokationspolaritet. De har kun fire meget konserverede sekvensmotiver og danner ringstrukturer eller ringe.
De blev først beskrevet i bakterier og bakteriofager. De er en gruppe replikerende eller replikative helikaser.
De har en unik 5'-3 'translokationspolaritet og har fem meget bevarede sekvensmotiver. Disse helikaser er kendetegnet ved dannelse af ringe.
De er proteiner af typen Rho-faktor. Helikaserne i SF5-superfamilien er karakteristiske for prokaryote organismer og er hexameriske ATP-afhængige. De menes at være tæt beslægtede med SF4; derudover har de ringformede og ikke-ringformede former.
De er proteiner, der tilsyneladende er beslægtede med SF3-superfamilien; SF6'er præsenterer imidlertid et domæne af ATPase-proteiner associeret med forskellige cellulære aktiviteter (AAA-proteiner), der ikke er til stede i SF3.
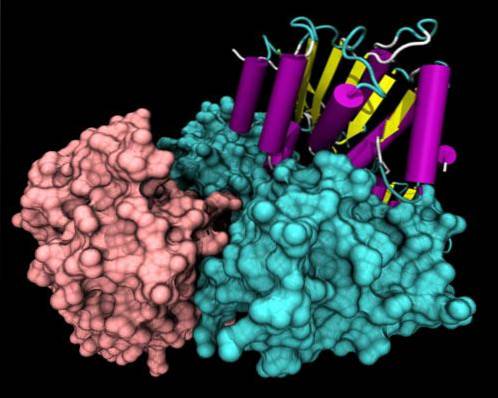
Strukturelt har alle helikaser meget konserverede sekvensmotiver i den forreste del af deres primære struktur. En del af molekylet har et bestemt aminosyrearrangement, der afhænger af den specifikke funktion af hver helicase..
De mest strukturelt studerede helikaser er SF1-superfamilien. Det er kendt, at disse proteiner er grupperet i 2 domæner, der ligner de multifunktionelle RecA-proteiner, og disse domæner danner en ATP-bindingslomme mellem dem..
Ikke-konserverede regioner kan præsentere specifikke domæner af DNA-genkendelsestypen, cellelokaliseringsdomænet og protein-protein..
Funktionerne af disse proteiner afhænger af en vigtig række forskellige faktorer, herunder miljømæssig stress, cellelinie, genetisk baggrund og cellecyklusfaser..
SF1 DNA-helikaser er kendt for at udføre specifikke funktioner i DNA-reparation, replikering, overførsel og rekombination..
De adskiller tråde af en DNA-dobbelthelix og deltager i vedligeholdelse af telomer, dobbeltstrengsreparation og fjernelse af nukleinsyreassocierede proteiner.
Som tidligere nævnt er RNA-helikaser vitale i langt størstedelen af RNA-metaboliske processer, og det er også kendt, at disse proteiner deltager i påvisningen af viralt RNA..
Derudover virker de i det antivirale immunrespons, da de detekterer fremmed RNA eller fremmed for kroppen (hos hvirveldyr).
Hellicases hjælper celler med at overvinde endogen og eksogen stress, undgår kromosomal ustabilitet og opretholder cellulær balance.
Svigt i dette system eller homeostatisk ligevægt er relateret til genetiske mutationer, der involverer gener, der koder for proteiner af helicase-typen; af denne grund er de genstand for biomedicinske og genetiske undersøgelser.
Nedenfor vil vi nævne nogle af sygdommene relateret til mutationer i gener, der koder DNA som proteiner af helicase-typen:
Det er en genetisk sygdom forårsaget af en mutation i et gen kaldet WRN, som koder for en helicase. Den mutante helikase fungerer ikke korrekt og forårsager en række sygdomme, der tilsammen udgør Werner syndrom.
Hovedkarakteristikken for dem, der lider af denne patologi, er deres for tidlige aldring. For at sygdommen skal manifestere sig, skal det mutante gen arves fra begge forældre; dens forekomst er meget lav, og der er ingen behandling for dens helbredelse.
Bloom syndrom er en genetisk sygdom, der skyldes mutationen af et autosomalt gen kaldet BLM, der koder for et helicase-protein. Det forekommer kun for personer, der er homozygote for den karakter (recessiv).
Hovedkarakteristikken ved denne sjældne sygdom er overfølsomhed over for sollys, som forårsager hudlæsioner af det erytromatøse udslæt. Der er ingen kur endnu.
Det er også kendt som medfødt atrofisk poikiloderma. Det er en patologi af meget sjælden genetisk oprindelse: til dato er der mindre end 300 tilfælde beskrevet over hele verden.
Det er forårsaget af en mutation i RECQ4-genet, et autosomalt og recessivt gen, der er placeret på kromosom 8.
Symptomer eller tilstande for dette syndrom inkluderer ungdommelig grå stær, abnormiteter i knoglesystemet, depigmentering, kapillær dilatationer og atrofi i huden (poikiloderma). I nogle tilfælde kan hyperthyreoidisme og mangel på testosteronproduktion forekomme..



Endnu ingen kommentarer