Det ontogeni det er den proces, hvor udviklingen af et individ finder sted. Fænomenet begynder med befrugtning og strækker sig til ældning af organiske væsener. Det biologiske felt med ansvar for at studere ontogeni er udviklingsbiologi.
I denne proces forekommer "oversættelsen" af genotypen - al den genetiske information fra en biologisk enhed - til den fænotype, som vi kan observere. Den mest dramatiske transformation sker i de tidlige stadier af udviklingen med transformation af en celle til et komplet individ.
I dag er sammensmeltningen af udviklingsbiologi og evolutionsteori, kendt som evo-devo, en meget populær viden, der vokser med stormskridt. Dette nye felt har til formål at forklare udviklingen af den enorme mangfoldighed af morfologier, der udstilles af levende organismer..
Artikelindeks
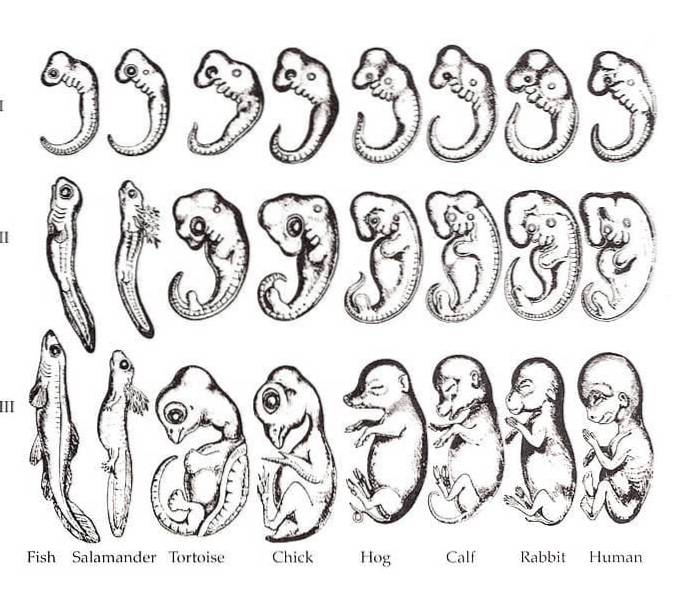
Forholdet mellem ontogeni og fylogeni var en fremherskende opfattelse gennem det 21. århundrede. Det er almindeligt kendt, at forskellige arter af organismer ligner hinanden meget mere i deres fosterstadier, end de er i voksne former. I året 1828 bemærkede Karl Ernst von Baer dette mønster i sybphylum Vertebrata.
Baer bemærkede, at der i forskellige arter af tetrapoder er visse ligheder i fosteret, såsom gæller, notokord, segmentering og finneformede lemmer..
Disse dannes før de typiske egenskaber, der gør det muligt for den pågældende gruppe at blive diagnosticeret i en mere specifik hierarkisk klassificeringsrækkefølge.
Denne idé blev omformuleret af den berømte - og en af Charles Darwins mest lidenskabelige tilhængere - tyskfødte biolog Ernst Haeckel..
Haeckel krediteres den berømte sætning "ontogeny recapitulates fylogeny." Med andre ord foreslår rekapitulation, at udviklingen af en organisme gentager dens evolutionære historie fra de voksne former for dens forfædre..
Selvom udtrykket er velkendt i dag, var det i midten af det 21. århundrede klart, at Haeckels forslag sjældent er opfyldt..
S. J. Gould, den berømte paleontolog og evolutionsbiolog, lagde sine ideer om rekapitulation i det, han kaldte "terminaladditionsprincippet." For Gould kan rekapitulation forekomme, så længe evolutionær ændring sker ved successiv tilføjelse af stadier i slutningen af en forfædres ontogeni..
På samme måde skulle det også opfyldes, at den tidlige varighed af forfædres ontogeni skulle afkortes, efterhånden som slægten udviklede sig.
I dag har moderne metoder formået at tilbagevise begrebet tilføjelse foreslået af den biogenetiske lov.
For Haeckel opstod denne tilføjelse på grund af den kontinuerlige anvendelse, der blev givet til organerne. Imidlertid er de evolutionære konsekvenser af brug og brug af organer blevet kasseret.
Det er i øjeblikket kendt, at gælbuerne i de embryonale faser hos pattedyr og krybdyr aldrig har den form, der svarer til voksne fisk.
Derudover er der variationer i timing eller timing af visse udviklingstrin. I evolutionær biologi kaldes denne ændring heterochrony..
Ontogeny omfatter alle processerne i udviklingen af organiske væsener, der begynder med befrugtning og slutter med aldring..
Logisk set forekommer de mest dramatiske transformationer i de første faser, hvor en enkelt celle er i stand til at danne et helt individ. Dernæst vil vi beskrive ontogeniprocessen og understrege de embryonale stadier.
Under processen med oogenese forbereder et æg (den kvindelige kønsceller, også kaldet et æg) til befrugtning og de tidlige stadier af udviklingen. Dette sker ved at akkumulere reservemateriale til fremtiden..
Æggets cytoplasma er et miljø rig på forskellige biomolekyler, hovedsageligt messenger-RNA, ribosomer, transfer-RNA og andre maskiner, der er nødvendige for syntesen af proteiner. Cellekernen gennemgår også betydelig vækst.
Sæd kræver ikke denne proces, deres strategi er at eliminere så meget cytoplasma som muligt og kondensere kernen for at bevare små dimensioner..
Den begivenhed, der markerer begyndelsen på ontogeni, er befrugtning, som involverer foreningen af en mandlig og kvindelig gamet, generelt under seksuel reproduktion..
I tilfælde af ekstern befrugtning, som forekommer i mange marine organismer, udvises begge kønsceller i vandet og findes tilfældigt.
Ved befrugtning integreres individets diploide antal igen og tillader kombinationsprocesser mellem de faderlige og moderens gener.
I visse tilfælde er sæd ikke nødvendigt for at aktivere udvikling. Men hos de fleste individer udvikler embryoet sig ikke på den rigtige måde. Tilsvarende kan nogle arter reproducere ved parthenogenese, hvor normal embryoudvikling forekommer uden behov for sæd..
I modsætning hertil kræver nogle æg aktivering af sædcellerne, men indarbejder ikke det genetiske materiale af denne mandlige gamet i fosteret..
Sæd og æg skal genkendes korrekt, så alle begivenheder efter befrugtning kan finde sted. Denne anerkendelse formidles af en række artsspecifikke proteiner. Der er også barrierer, der forhindrer, at et æg, når det først er befrugtet, nås af en anden sæd.
Efter befrugtning og aktivering af ægget forekommer de første udviklingsstadier. I segmentering deler embryoet sig gentagne gange for at blive en gruppe celler kaldet blastomerer..
I denne sidste periode forekommer der ingen cellevækst, kun underopdelingen af massen finder sted. I sidste ende er der hundreder eller tusinder af celler, der giver plads til blastula-staten.
Når embryoet udvikler sig, får det en polaritet. Af denne grund er det muligt at skelne mellem plantepolen, der er placeret i den ene ende, og dyrepolen, der er rig på cytoplasma. Denne akse er et referencepunkt for udvikling.
Afhængig af mængden af æggeblomme, som ægget har, og fordelingen af stoffet, kan ægget klassificeres som oligolecytter, heterolecytter, telolecytter og centrolecytter..
Førstnævnte har, som navnet antyder, en lille mængde æggeblomme, og den er mere eller mindre jævnt fordelt over ægget. Generelt er størrelsen lille. Heterolecytter har mere æggeblomme end oligolecytter, og æggeblomme er koncentreret i den vegetative pol.
Telolecitos har en rigelig mængde æggeblomme, der optager næsten hele æg. Endelig har centrolecitos al æggeblommen koncentreret i æggets centrale region..
Blastula er en masse celler. Hos pattedyr kaldes denne cellegruppe en blastocyst, mens cellerne i de fleste dyr er arrangeret omkring et centralt væskehulrum, kaldet en blastocele..
I blastula-tilstand har det været muligt at vise en stor stigning med hensyn til mængden af DNA. Imidlertid er størrelsen på hele fosteret ikke meget større end den oprindelige zygote..
Gastrulering omdanner den sfæriske og enkle blastula til en meget mere kompleks struktur med to kimlag. Denne proces er heterogen, hvis vi sammenligner de forskellige slægter af dyr. I nogle tilfælde dannes et andet lag uden at skabe et indre hulrum.
Åbningen til tarmen kaldes blastopore. Blastopores skæbne er et meget vigtigt kendetegn for opdelingen af to store slægter: protostomaterne og deuterostomes. I den første gruppe giver blastopore munden, mens i den anden blastopore giver anus.
Således har gastrula to lag: et ydre lag, der omgiver blastocele, kaldet ektoderm, og et indre lag kaldet endoderm..
De fleste dyr har et tredje kimlag, mesoderm, placeret mellem de to ovennævnte lag. Mesoderm kan dannes på to måder: cellerne stammer fra en ventral region af blastopore-læben, og derfra spreder de sig eller opstår fra den centrale region af archnteronvæggene..
I slutningen af gastrulation dækker ektoderm embryoet, og mesoderm og endoderm er placeret i den indre del. Med andre ord har cellerne en anden slutposition end den, hvor de startede..
Coelom er et kropshulrum, der er omgivet af mesoderm. Dette sker, fordi blastocele under gastrulationsprocessen næsten er fuldstændigt fyldt med mesoderm..
Dette coelomatiske hulrum kan vises på to måder: schizocelic eller enterocelic. Imidlertid er begge coeloms funktionelt ækvivalente.
Organogenese omfatter en række processer, hvor hvert af organerne dannes.
De mest relevante begivenheder inkluderer migration af bestemte celler til det sted, hvor de er nødvendige for at danne nævnte organ..
Under udvikling er det blevet bestemt, at epigenesen fortsætter i tre faser: mønsterdannelse, bestemmelse af kropsposition og induktion af den korrekte position for ekstremiteterne og forskellige organer.
For at generere et respons er der visse genprodukter, kaldet morfogener (definitionen af disse enheder er teoretisk, ikke kemisk). Disse fungerer takket være dannelsen af en differentiel gradient, der leverer rumlig information.
Med hensyn til de involverede gener spiller homeotiske gener en grundlæggende rolle i individernes udvikling, da de definerer identiteten af segmenterne.



Endnu ingen kommentarer