Det lov om økologisk tiende, økologisk lovgivning eller 10% hæver den måde, hvorpå energi bevæger sig i dens afledning gennem de forskellige trofiske niveauer. Det hævdes også ofte, at denne lov simpelthen er en direkte konsekvens af den anden lov om termodynamik.
Økologisk energi er en del af økologien, der beskæftiger sig med at kvantificere de forhold, som vi har skitseret ovenfor. Det anses for, at Raymond Lindemann (specifikt i hans banebrydende arbejde i 1942), var den, der grundlagde grundlaget for dette studieområde.
Hans arbejde fokuserede på begreberne fødekæde og web og på kvantificering af effektiviteten i overførsel af energi mellem de forskellige trofiske niveauer..
Lindemann starter fra den indfaldende solstråling eller energi, som et samfund modtager, gennem fangsten udført af planter gennem fotosyntese og fortsætter med at overvåge denne fangst og dens efterfølgende anvendelse af planteædere (primære forbrugere), derefter af kødædere (sekundære forbrugere) og endelig af nedbrydere.
Artikelindeks
Efter Lindemanns banebrydende arbejde blev trofisk overførselseffektivitet antaget at være omkring 10%; faktisk henviste nogle økologer til en lov på 10%. Siden da er der imidlertid genereret flere forvirringer vedrørende dette problem..
Der er bestemt ingen naturlov, der resulterer i, at nøjagtigt en tiendedel af energien, der kommer ind i det ene trofiske niveau, overføres til det næste..
For eksempel afslørede en samling trofiske undersøgelser (i hav- og ferskvandsmiljøer), at overførselseffektiviteter efter trofisk niveau lå mellem ca. 2 og 24%, skønt gennemsnittet var 10,13%..
Som en generel regel, der gælder for både vand- og terrestriske systemer, kan det siges, at den sekundære produktivitet fra planteædere normalt er placeret omtrent, en størrelsesorden under den primære produktivitet, som den er baseret på..
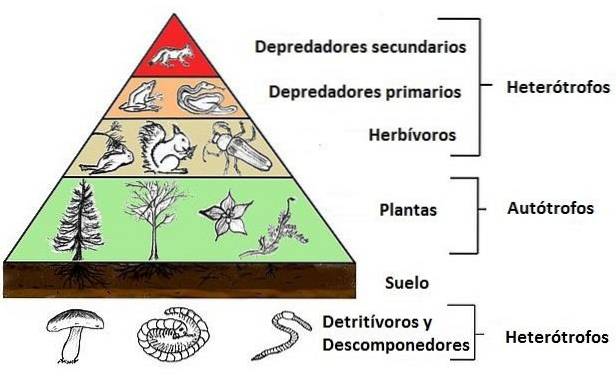
Dette er ofte et konsistent forhold, der opretholdes i alle foderingssystemer, og som har tendens til at blive strukturer af pyramidetype, hvor basen leveres af planterne, og på denne base er en mindre baseret, af de primære forbrugere, hvorpå en anden (endnu mindre) sekundære forbrugere sidder.
Alle levende ting kræver materie og energi; sager for opbygningen af deres kroppe og energi til at udføre deres vitale funktioner. Dette krav er ikke begrænset til en individuel organisme, men det udvides til højere niveauer af biologisk organisation, som disse individer kan overholde..
Disse organisationsniveauer er:
I et økosystem skaber samfundet og miljøet strømme af energi og stof.
Organismerne i et økosystem er grupperet efter en "rolle" eller "funktion", som de udfører inden for fødevarer eller trofiske kæder; det er sådan, vi taler om trofiske niveauer hos producenter, forbrugere og nedbrydere.
Til gengæld interagerer hvert eneste af disse trofiske niveauer med det fysisk-kemiske miljø, der giver livsvilkårene og på samme tid fungerer som en kilde og synke af energi og stof..
For det første skal vi definere den primære produktivitet, som er den hastighed, hvormed der produceres biomasse pr. Arealenhed..
Det udtrykkes normalt i energienheder (Joule pr. Kvadratmeter pr. Dag) eller i enheder af tørt organisk stof (kg pr. Hektar og pr. År) eller som kulstof (masse af kulstof i kg pr. Kvadratmeter pr. År).
Generelt, når vi henviser til al den energi, der er fastlagt ved fotosyntese, kalder vi det normalt bruttoprimærproduktivitet (PPG).
Heraf bruges en andel i åndedrættet af de samme autotrofer (RA) og går tabt i form af varme. Nettoprimærproduktionen (PPN) opnås ved at trække denne mængde fra PPG (PPN = PPG-RA).
Denne primære nettoproduktion (PPN) er det, der i sidste ende er tilgængeligt til forbrug af heterotrofer (disse er bakterier, svampe og resten af de dyr, som vi kender).
Sekundær produktivitet (PS) er defineret som hastigheden af ny biomasseproduktion af heterotrofiske organismer. I modsætning til planter, heterotrofe bakterier, svampe og dyr kan de ikke fremstille de komplekse og energirige forbindelser, de har brug for, fra simple molekyler..
De får altid deres stof og energi fra planter, som de kan gøre direkte ved at forbruge plantemateriale eller indirekte ved at fodre på andre heterotrofer..
Det er på denne måde, at planter eller fotosyntetiske organismer generelt (også kaldet producenter) udgør det første trofiske niveau i et samfund; primære forbrugere (dem der lever af producenter) udgør det andet trofiske niveau og sekundære forbrugere (også kaldet kødædere) udgør det tredje niveau.
Proportionerne af nettoprimærproduktion, der strømmer langs hver af de mulige energibaner, afhænger i sidste ende af overførselseffektiviteter, det vil sige på den måde, hvorpå energi bruges og overføres fra et niveau til et andet..
Der er tre kategorier af energioverførselseffektivitet, og med disse veldefinerede kan vi forudsige mønsteret for energistrøm på trofiske niveauer. Disse kategorier er: forbrugseffektivitet (EF), assimileringseffektivitet (EA) og produktionseffektivitet (EP).
Lad os nu definere disse tre nævnte kategorier.
Matematisk kan vi definere forbrugseffektiviteten (EF) som følger:
EC =jegn/Pn-1 × 100
Hvor vi kan se, at CE er en procentdel af den samlede tilgængelige produktivitet (Pn-1), der effektivt indtages af det øvre sammenhængende trofiske rum (jegn).
For primære forbrugere i græsningssystemet er EF f.eks. Den procentdel (udtrykt i energienheder og pr. Tidsenhed) af PPN, der forbruges af planteædere.
Hvis vi henviste til sekundære forbrugere, ville det svare til procentdelen af produktiviteten af planteædere, der forbruges af kødædere. Resten dør ikke spist og går ind i forfaldskæden.
På den anden side udtrykkes assimileringseffektiviteten som følger:
EA =TILn/jegn × 100
Igen henviser vi til en procentdel, men denne gang til den del af energien, der kommer fra maden og indtages i et trofisk rum af en forbruger (jegn) og det assimileres af dit fordøjelsessystem (TILn).
Denne energi vil være den, der er tilgængelig til vækst og til udførelse af arbejde. Resten (den del, der ikke er assimileret), går tabt med fæces og går derefter ned på dekomponenternes trofiske niveau.
Endelig udtrykkes produktionseffektiviteten (EP) som:
EP = Pn/TILn × 100
hvilket også er en procentdel, men i dette tilfælde henviser vi til den assimilerede energi (TILn) der ender med at blive inkorporeret i ny biomasse (Pn). Al den uassimilerede energirest går tabt i form af varme under respiration.
Produkter som sekreter og / eller udskillelser (rig på energi), som har deltaget i metaboliske processer, kan betragtes som produktion, Pn, og er tilgængelige, ligesom lig, for nedbrydere.
Efter at have defineret disse tre vigtige kategorier kan vi nu spørge os selv om den "globale overførselseffektivitet" fra et trofisk niveau til det næste, som simpelthen er givet af produktet af de ovennævnte effektivitetsgevinster (EC x EA x EP).
Udtrykt i daglig tale kan vi sige, at effektiviteten af et niveau er givet af det, der effektivt kan indtages, som derefter assimileres og ender med at blive inkorporeret i ny biomasse.
Planteædende produktivitet er altid lavere end for planterne, som de fodrer med. Vi kunne så spørge os selv: Hvor går den spildte energi hen?
For at besvare dette spørgsmål skal vi henlede opmærksomheden på følgende fakta:
Dette sker af to grundlæggende grunde: For det første på grund af det faktum, at der ikke er nogen energiomdannelsesproces, der er 100% effektiv. Det vil sige, at der altid er et tab i form af varme i omdannelsen, hvilket er i fuld overensstemmelse med den anden lov om termodynamik..
For det andet, da dyr har brug for at arbejde, hvilket kræver energiforbrug, og det igen medfører nye tab i form af varme..
Disse mønstre forekommer på alle trofiske niveauer, og som forudsagt af anden lov om termodynamik spredes en del af den energi, som man forsøger at overføre fra et niveau til et andet, altid i form af ubrugelig varme..



Endnu ingen kommentarer