
Det genetisk rekombination det er den proces, hvormed nukleinsyremolekyler udveksler fragmenter og genererer et nyt molekyle. Det er meget almindeligt i DNA, men RNA er også et substrat til rekombination. Rekombination er efter mutation den vigtigste kilde til generering af genetisk variabilitet.
DNA deltager i forskellige biokemiske processer. Under replikering fungerer det som en skabelon til generering af to nye DNA-molekyler. I transkription tillader det dannelse af RNA-molekyler fra specifikke regioner kontrolleret af en promotor.
Men derudover er DNA også i stand til at udveksle fragmenter. Gennem denne proces genererer det nye kombinationer, der ikke er et produkt af de to foregående processer eller af befrugtning..
Al rekombinationsproces involverer nedbrydning og sammenføjning af de DNA-molekyler, der deltager i processen. Denne mekanisme varierer afhængigt af rekombinationssubstratet, de enzymer, der deltager i processen, og mekanismen til dets udførelse..
Rekombination afhænger generelt af eksistensen af komplementære, ens (hvis ikke identiske) eller homologe regioner mellem de rekombinerende molekyler. I tilfælde af at molekyler rekombineres i processer, der ikke styres af homologi, siges rekombinationen at være ikke-homolog.
Hvis homologien involverer en meget kort region til stede i begge molekyler, siges rekombinationen at være stedsspecifik..
Artikelindeks
Det, vi kalder homologi i rekombination, refererer ikke nødvendigvis til de deltagende molekylers evolutionære oprindelse. Vi taler mere om graden af lighed i nukleotidsekvensen.
Ikke-reparativ rekombination forekommer for eksempel i eukaryoter under meiose-processen. Der kan utvivlsomt ikke være større homologi end mellem par af kromosomer i samme celle.
Derfor kaldes de homologe kromosomer. Der er dog tilfælde, hvor DNA fra en celle udveksler materiale med et fremmed DNA. Disse DNA'er skal være meget lig rekombination, men de behøver ikke nødvendigvis at dele den samme forfader (homologi) for at opnå det..
Stedet for vedhæftning og udveksling mellem to DNA-molekyler kaldes chiasme, og selve processen kaldes tværbinding. Under overgangen verificeres en udveksling af bånd mellem de deltagende DNA'er..
Dette genererer et møntintegrat, som er to DNA-molekyler, der fysisk er forbundet til et. Ved "løsning" af møntintegreringen genereres to molekyler, generelt ændrede (rekombinant).
At "løse" i forbindelse med rekombination er at adskille komponent-DNA-molekylerne i et møntintegrat.
I stedsspecifik rekombination har to DNA-molekyler, generelt ikke homologe, en kort sekvens, der er fælles for begge. Denne sekvens er målet for et specifikt splejsningsenzym.
Enzymet, der er i stand til at genkende denne sekvens og ikke en anden, skærer det på et bestemt sted i begge molekyler. Ved hjælp af nogle andre faktorer udveksler det DNA-båndene i de to deltagende molekyler og danner et møntintegrat.
Dette er grundlaget for dannelsen af møntintegreringen mellem bakteriens genom Escherichia coli og bakteriofag lambda. En bakteriofag er en virus, der inficerer bakterier.
Dannelsen af dette møntintegrat udføres af et enzym kodet i virusgenomet: lambda-integrase. Den genkender en fælles sekvens kaldet attP i virusets cirkulære genom og attB i bakterienes.
Ved at skære begge sekvenser i begge molekyler genererer det frie segmenter, bytter båndene og forbinder de to genomer. Derefter dannes en større eller møntintegreret cirkel.
I møntintegration bæres virusgenomet passivt af det bakterielle genom, som det replikerer med. I denne tilstand siges det, at virussen er i provirus-tilstand, og at bakterien er lysogen for den..
Den omvendte proces, det vil sige opløsning af møntintegreringen, kan tage mange generationer - eller endda ikke forekomme. Imidlertid medieres dette enzymatisk af et andet protein kodet af virusgenomet kaldet excisionase. Når dette sker, adskilles virussen fra cointegratet, genaktiveres og forårsager cellelyse..
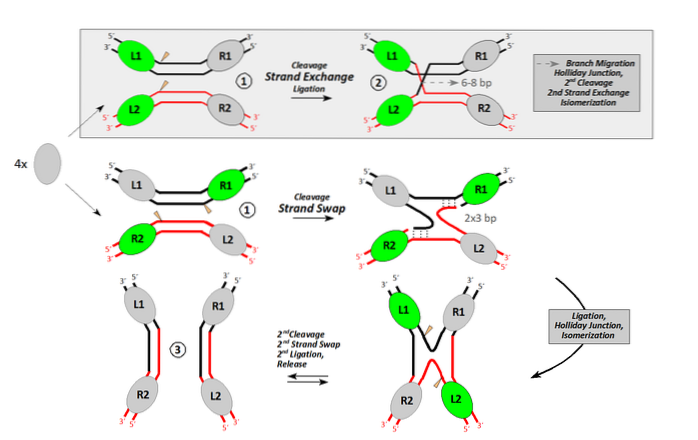
Homolog rekombination finder sted mellem DNA-molekyler, der deler mindst ca. 40 nukleotider med fuldstændig eller næsten fuldstændig lighed. For at udføre rekombinationsprocessen skal mindst en endonuklease deltage.
Endonukleaser er enzymer, der foretager interne nedskæringer i DNA. Nogle gør det for at fortsætte med at nedbryde DNA. Andre, som i tilfælde af rekombination, gør det for at generere en buler i DNA'et.
Dette unikke nick tillader behandling af et enkelt bånd DNA med en fri ende. Denne frie ende, orienteret af en rekombinase, gør det muligt for et enkelt bånd at invadere et dobbelt DNA, der fortrænger det residente bånd identisk med det..
Dette er krydsningspunktet mellem en donor ("invader") DNA-molekyle og en anden receptor..
Enzymet (rekombinase), der udfører processen med båndinvasion og udveksling Escherichia coli det kaldes RecA. Der er andre homologe proteiner i prokaryoter, såsom RadA i archaea. I eukaryoter kaldes det ækvivalente enzym RAD51.
Når det invasive bånd fortrænger beboeren, interagerer hun med det bånd, der blev efterladt simpelt i donormolekylet. Begge steder forsegles ved indvirkning af en ligase.
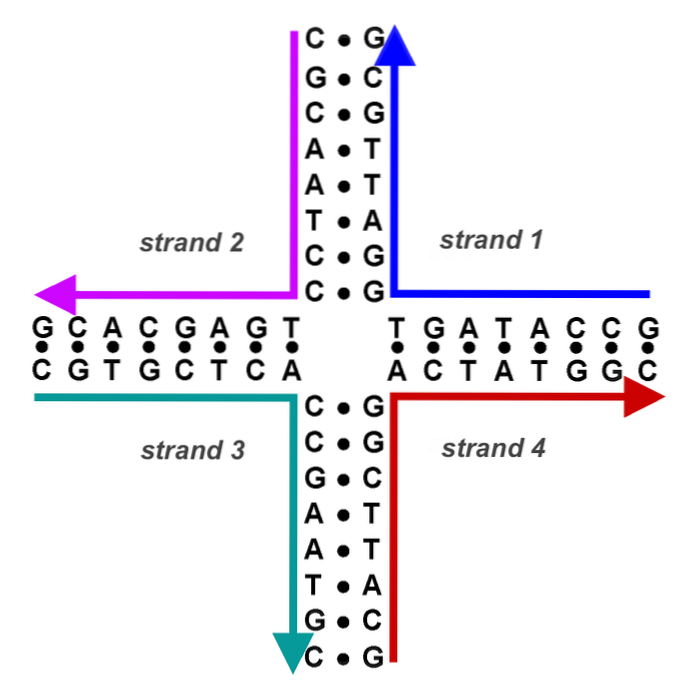
Nu har vi et DNA af hybridbånd (et donorbånd og et modtagerbånd af forskellig oprindelse) flankeret af donor-DNA og modtager-DNA. Crossover-punkterne (chiasmata) bevæger sig i begge retninger med mindst 200 bp.
Hvert punkt af crossover danner såkaldt Holliday-strukturen (korsformet DNA-produkt fra en rekombinationshændelse).
Dette korsformede DNA skal løses af andre endonukleaser. Kimærisk eller hybrid DNA med denne struktur kan løses på to måder. Hvis den anden endonukleotidsnit sker i samme bånd som den første, sker der ingen rekombination. Hvis det andet snit forekommer i det andet bånd, er de resulterende produkter rekombinante.
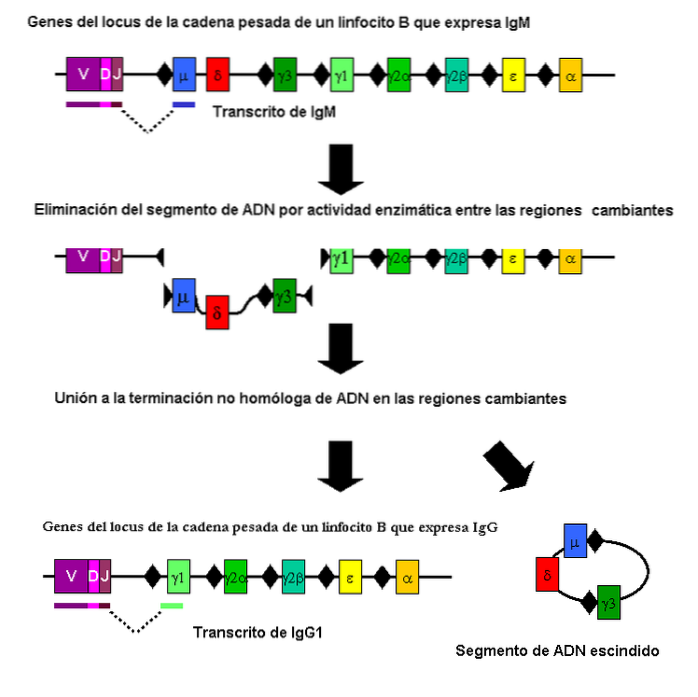
Dette er en type somatisk rekombination (ikke meiotisk), der bidrager til dannelsen i den enorme variation i antistoffer mod immunsystemet.
Denne rekombination finder sted i særlige fragmenter af generne, der koder for de polypeptidkæder, der definerer dem. Det udføres af B-celler og involverer forskellige genetiske regioner.
Interessant er der parasitter som Trypanosoma brucei der anvender en lignende rekombinationsmekanisme til at skabe variation i et overfladeantigen. På denne måde kan de unddrage sig værtens respons, hvis værten ikke genererer det antistof, der er i stand til at genkende det "nye" antigen..
Endelig er der rekombinationsprocesser, der ikke afhænger af ligheden i sekvensen af de deltagende molekyler. I eukaryoter er f.eks. Rekombinationen af ikke-homologe ender meget vigtig.
Dette sker med DNA-fragmenter, der viser dobbeltbåndsbrud i DNA'et. Disse "repareres" ved at cellen forbinder dem med andre fragmenter også med dobbeltbåndsbrud.
Disse molekyler behøver imidlertid ikke nødvendigvis at være ens for at deltage i denne rekombinationsproces. Ved at reparere skaden kan cellen slutte sig til ikke-relaterede DNA'er og derved skabe et virkelig nyt (rekombinant) molekyle.
Rekombination sikrer troskab af DNA-information under og efter replikationsprocessen. Rekombination registrerer DNA-beskadigelse under den nye båndproces i dette ekstremt lange makromolekyle.
Da hvert bånd har sine egne oplysninger, og dets komplementære, garanterer rekombination, at ingen går tabt. Hver optræder som et vidne for den anden. Tilsvarende i diploide organismer er et homologt kromosom vidne til dets søskende og omvendt.
På den anden side varierer skadereparationsmekanismerne for dette molekyle, når DNA er blevet replikeret. Nogle er direkte (skaden handles direkte), og andre er indirekte.
Indirekte reparationsmekanismer afhænger af, at rekombination finder sted. Det vil sige at der bruges et andet homologt molekyle til at reparere skaden i et DNA-molekyle. Dette ville fungere i reparativ rekombination som en skabelon, hvorfra den har lidt skade..
Rekombination er i stand til at skabe enorm kromosomal variation under meiose. Somatisk rekombination genererer også variation, som i tilfælde af antistoffer hos hvirveldyr.
I mange organismer er meiose gametisk. I seksuelt reproducerede organismer er rekombination en af de mest kraftfulde måder at generere variation på.
Det vil sige, at til spontan mutation og kromosomseparation skal rekombination tilføjes som et andet element, der genererer gametisk variation..
Integrationen af bakteriofaggenomer ved stedsspecifik rekombination har på den anden side bidraget til ombygning af genomet af deres værtsbakterier..
Dette har bidraget til dannelsen af genomisk variation og udvikling af denne vigtige gruppe af levende væsener..
Vi har allerede set, at DNA kan repareres, men ikke hvad der skader det. I virkeligheden kan næsten alt skade DNA, startende med defekt replikering, der ikke korrigeres..
Men ud over det kan DNA blive beskadiget af UV-lys, ioniserende stråling, frie iltradikaler fra cellulær respiration, og hvad vi spiser, ryger, indånder, indtager eller berører..
Heldigvis behøver du ikke opgive at leve for at beskytte DNA. Visse ting skal opgives, men det store arbejde udføres af cellen selv. Disse mekanismer til påvisning af beskadigelse af DNA og dets reparation har naturligvis et genetisk grundlag og deres mangel enorme konsekvenser.
Sygdomme relateret til defekter i homolog rekombination indbefatter for eksempel Bloom- og Werner-syndromer, familiær bryst- og æggestokkræft osv..



Endnu ingen kommentarer