Det celleblære Det er et middel til intracellulær og ekstracellulær kommunikation, hvor molekyler syntetiseret i cellen, såsom neurotransmittere, hormoner, proteiner, lipider og nukleinsyrer, er pakket. Disse molekyler kaldes fragt. Opladningens kemiske natur afhænger af typen af galdeblæren og dens funktion.
En vesikels generelle morfologi består af et lipiddobbeltlag, der danner en lukket sæk, og hvis lumen er vandig. Størrelsen på vesiklerne kan variere. For eksempel i bugspytkirtlens acinære celler varierer det fra 200 til 1200 nm, mens det i neuroner varierer fra 30 til 50 nm.
I eukaryoter forekommer forskellige cellulære processer i specifikke organeller. Det er dog nødvendigt at udveksle molekyler mellem organeller eller at sende molekyler ud i det ekstracellulære rum. På grund af dette er der behov for et system, der gør det muligt at transportere lasten til sin korrekte destination. Denne funktion er opfyldt af vesiklerne.
Artikelindeks
Der findes forskellige typer vesikulær transport med deres respektive egenskaber. Imidlertid er der generaliteter såsom spiring, som er rettet af et lag eller overtrukket med proteiner, såsom clathrin; og bindingsspecificitet, som afhænger af transmembranproteiner eller SNARE.
Vesikulær transport inkluderer exocytose og endocytose, transport mellem organeller og frigivelse af ekstracellulære vesikler. I alle tilfælde involverer det kontinuerlig dannelse af skud og spaltning og fusion af transportvesikler..
Eksocytose består af fusion af en vesikel med plasmamembranen for at frigive vesikulært indhold. Der er tre former for eksocytose: 1) fuldstændig kollapsfusion; 2) kys og løb; og 3) sammensat eksocytose.
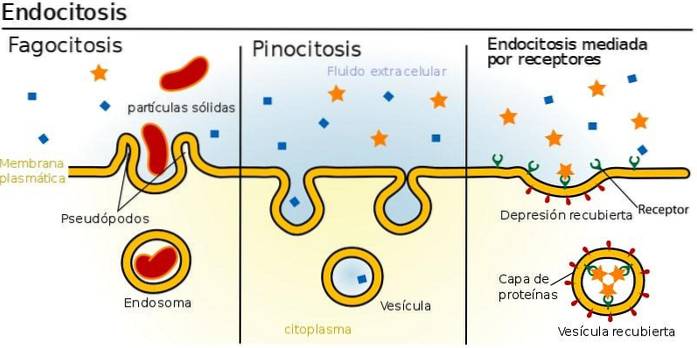
Endocytose består af inddrivelsen af plasmamembranen, som forhindrer cellulær betændelse. Der er forskellige mekanismer for endocytose.
I vesikulær transport mellem organeller transporteres de nyligt syntetiserede proteiner, der findes i lumen i det endoplasmatiske retikulum, til Golgi-apparatet. Fra denne organelle afgår vesiklerne mod endomembransystemet og plasmamembranen..
Ekstracellulære vesikler, der findes i prokaryoter og eukaryoter, er ansvarlige for at bære molekyler fra en celle til en anden.
De tjener til at introducere molekyler i cellen eller til at genbruge membrankomponenter. Disse vesikler er måske ikke dækket af et lag af proteiner. Proteinerne, der beklæder overfladen af galdeblæren, er clathrin og caveolin..
Clathrin-coatede endocytiske vesikler er ansvarlige for internaliseringen af patogener, såsom influenzavirus blandt andre, membranproteiner og ekstracellulære receptorer og ligander. Caveolin-coatede vesikler medierer indtrængen af vira, svampe, bakterier og prioner.
Gennem en stimulus frigiver sekretoriske celler (neuroner eller andre celler) deres indhold gennem exocytocis.
Membranfusion under exocytose forekommer gennem to trin: 1) binding af den exocytiske vesikel til membranacceptoren; og 2) fusion af lipiddobbeltlagene. Rab-, GTPases- og SNARE-proteinerne deltager blandt andre i disse trin..
COPII-overtrukne vesikler transporteres fra det endoplasmatiske retikulum til Golgi-apparatet. Transport fra Golgi-apparatet til vakuolen involverer to veje: ALP (alkalisk phosphatase) til vakuolen; endosomer via carboxypeptidase Y- og S-vejen (CPY og CPS).
Blærerne i den sekretoriske vej har en bred vifte af funktioner, blandt hvilke sekretionen af følgende stoffer er: insulin fra bugspytkirtelceller, neuropeptider og neurotransmittere, hormoner og stoffer involveret i immunresponset.
En af de bedst kendte funktioner er frigivelsen af sekretoriske proteiner fra bugspytkirtlen. For eksempel frigives chymotrypsinogen, et zymogen, ved fusion af vesikler til membranen som et resultat af hormonstimulering..
Ekstracellulære vesikler (EV) er af to typer: exosomer og ektosomer. Begge er differentieret ved deres sammensætning, der bestemmer deres funktion. Eksosomer besidder tetraspanin, integrin, proteoglycan og ICAMI. Ektosomer har receptorer, glycoproteiner, metalloproteiner og nukleinsyrer.
EV'ernes funktioner inkluderer vedligeholdelse af cellehomeostase, regulering af cellefunktion og intercellulær kommunikation. Sidstnævnte funktion kræver transport af proteiner, RNA (mRNA, miRNA, ikke-kodende RNA) og DNA-sekvenser..
Fusion af EV'er til målcellemembranen kan påvirke reguleringen af genekspression ved transkriptionsfaktorer, signalproteiner og mange enzymer. EV'er frigivet af stamceller spiller en vigtig rolle i orgelreparation og beskyttelse mod sygdom.
Den normale fysiologiske funktion af celler afhænger blandt flere faktorer af transport af vesikler og deres fusion. F.eks. Er type 2-diabetes karakteriseret ved defekter i insulinsekretion og translokation medieret af glukosetransportører..
EV'er spiller en vigtig rolle i mange sygdomme. I kræft øger EV'er resistens over for kemoterapeutiske lægemidler, medieret af miRNA,
EV'er har en kritisk effekt på neurodegeneration. I Alzheimers sygdomme og multipel sklerose afhænger den degenerative effekt af flere molekyler, såsom miRNA, gangliosider og proteiner..
I hjerteceller tillader exosomer og ektosomer kommunikation mellem celler og påvirker også udviklingen af ateroklerotisk plaque i kar ved at inducere inflammation, spredning, trombose og vasoaktiv respons..
I allergi- og betændelsesprocesser regulerer miRNA'er af EV'er disse processer gennem parakrine effekter.
Særlig opmærksomhed er blevet brugt på EV'er af protozoer. Dette skyldes, at elbiler har en vigtig rolle mellem parasit og værtsinteraktion..
Nogle af de parasitter, hvis VE er blevet undersøgt, er Trypanosoma brucei, Trypanosoma cruzi, Leishmania spp., Plasmodium spp. og Toxoplasma spp.
EV'er er også blevet observeret i gram-positive og negative bakterier, herunder Corynebacterium og Moraxellaceae. I slimhinden i luftvejene binder ydre membranvesikler (OMV'er) til lipiddomæner i alveolære epitelceller. Derfra modulerer OMV det inflammatoriske respons.


Endnu ingen kommentarer