Det axoneme Det er en intern cytoskeletal struktur af cilia og flagella baseret på mikrotubuli, og som giver dem bevægelse. Dens struktur består af en plasmamembran, der omgiver et par centrale mikrotubuli og ni par perifere mikrotubuli..
Axoneme er placeret uden for cellen og er forankret inde i cellen ved hjælp af basallegemet. Den er 0,2 um i diameter, og dens længde kan variere fra 5-10 um i cilier til flere mm i flagellum af nogle arter, selvom disse generelt måler 50-150 um.
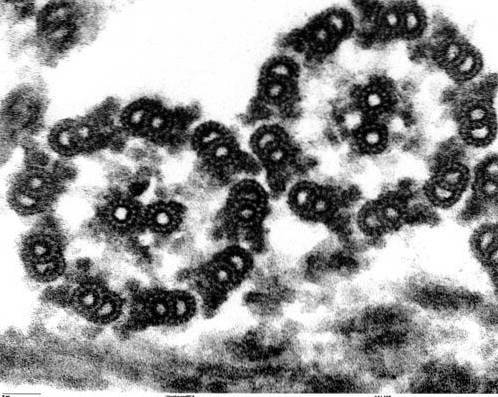
Axonemstrukturen af cilia og flagella er yderst konservativ i alle eukaryote organismer, fra mikroalger Chlamydomonas til plagen af den menneskelige sæd.
Artikelindeks
Aksonemerne hos langt størstedelen af cilier og flageller har en konfiguration kendt som "9 + 2", det vil sige ni par perifere mikrotubuli, der omgiver et centralt par.
Mikrotubuli i hvert par er forskellige i størrelse og sammensætning bortset fra det centrale par, der præsenterer begge mikrotubuli ens. Disse rør er stabile strukturer, der er i stand til at modstå brud..
Mikrotubuli har polaritet og har alle det samme arrangement, hvor deres "+" ende er placeret mod toppen og "-" enden er basalt..
Som vi allerede har påpeget, er aksonemets struktur af type 9 + 2. Mikrotubuli er lange cylindriske strukturer, der består af protofilamenter. Protofilamenter består til gengæld af proteinunderenheder kaldet alfa tubulin og beta tubulin..
Hver protofilament har en alfa-tubulin-enhed i den ene ende, mens den anden ende har en beta-tubulin-enhed. Enden med beta-tubulinterminalen kaldes "+" -enden, den anden ende ville være "-" -enden. Alle protofilamenter i den samme mikrotubuli er orienteret med den samme polaritet.
Mikrotubuli indeholder ud over tubuliner også proteiner kaldet mikrotubuli-relaterede proteiner (MAP). Af hvert par perifere mikrotubuli består den mindste (mikrotubuli A) af 13 protofilamenter.
Mikrotubuli B har kun 10 protofilamenter, men det er større end mikrotubuli A. Det centrale par mikrotubuli har samme størrelse, og hver af dem består af 13 protofilamenter.
Dette centrale par mikrotubuli er omsluttet af den centrale kappe, protein i naturen, som vil forbinde med de perifere A-mikrotubuli ved hjælp af de radiale stråler. På den anden side er mikrotubuli A og B i hvert par forbundet sammen med et protein kaldet nexin..
Mikrotubuli En del også et par arme dannet af et protein kaldet dynein. Dette protein er ansvarlig for at bruge den tilgængelige energi i ATP til at opnå bevægelse af cilia og flagella.
Eksternt er axonemet dækket af en ciliær eller flagellær membran, der har samme struktur og sammensætning som plasmamembranen i cellen..
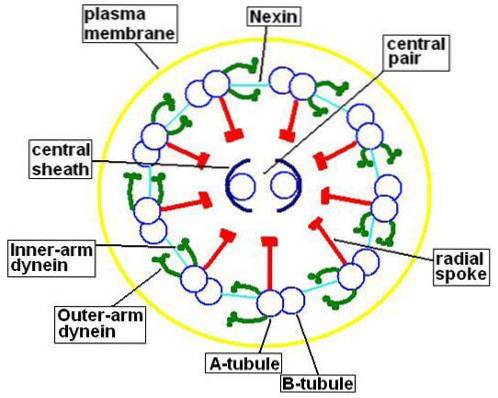
Selvom "9 + 2" -sammensætningen af axoneme er meget konserveret i de fleste eukaryote cilierede og / eller flagellerede celler, er der nogle undtagelser fra denne model..
I sæd fra nogle arter går det centrale par mikrotubuli tabt, hvilket resulterer i en "9 + 0" konfiguration. Flagellarbevægelsen i disse sædceller ser ikke ud til at variere meget fra den, der observeres i axonemer med normal konfiguration, for hvilket det antages, at disse mikrotubuli ikke spiller en vigtig rolle i bevægelsen..
Dette aksonemmønster er blevet observeret i sædceller fra arter såsom fisk Lycondontis og af annelider af slægten Myzostomum.
En anden konfiguration observeret i axonemes er “9 + 1” konfigurationen. I dette tilfælde er der en enkelt central mikrotubuli til stede snarere end et par. I sådanne tilfælde modificeres den centrale mikrotubuli grundigt og præsenterer adskillige koncentriske vægge.
Dette aksonem-mønster er blevet observeret i de mandlige kønsceller hos nogle arter af fladorm. I disse arter gentages dette axonememønster imidlertid ikke i andre flagellerede eller cilierede celler af organismer..
Undersøgelser af flagellabevægelse har vist, at flagellafleksion forekommer uden sammentrækning eller afkortning af mikrotubuli i axoneme. På grund af dette har cytologen Peter Satir foreslået en model for flagellær bevægelse baseret på forskydning af mikrotubuli..
Ifølge denne model opnås bevægelse takket være forskydningen af en mikrotubuli fra hvert par på sin partner. Dette mønster svarer til glidningen af myosinkæder på actin under muskelsammentrækning. Bevægelse sker i nærværelse af ATP.
Dyneinarmene er forankret i mikrotubuli A i hvert par, med enderne rettet mod mikrotubuli B. Ved begyndelsen af bevægelse klæber dyneinarmene til bindingsstedet på mikrotubuli B. Derefter sker der en ændring i konfigurationen af dyneinet, som driver mikrorør B nedad.
Nexin holder begge mikrotubuli tæt på hinanden. Derefter adskilles dyneinarmene fra mikrotubuli B. Derefter tilslutter det sig at gentage processen. Denne glidning sker skiftevis mellem den ene side af axoneme og den anden..
Denne skiftende forskydning på den ene side af axoneme får cilium eller flagellum til at bøje sig først til den ene side og derefter til den modsatte side. Fordelen ved Satirs flagellære bevægelsesmodel er, at det ville forklare appendiksens bevægelse uafhængigt af axonem-mikrotubuli's axonemkonfiguration.
Der er flere genetiske mutationer, der kan forårsage unormal udvikling af axoneme. Disse abnormiteter kan blandt andet være manglen på en af dyneinarmene, enten interne eller eksterne, i de centrale mikrotubuli eller de radiale stråler..
I disse tilfælde udvikles et syndrom kaldet Kartageners syndrom, hvor mennesker, der lider af det, er infertile, fordi sædcellerne ikke er i stand til at bevæge sig.
Disse patienter udvikler også indvolde i en omvendt position i forhold til den normale position; for eksempel hjertet placeret på højre side af kroppen og leveren til venstre. Denne tilstand er kendt som situs inversus.
Dem med Kartageners syndrom er også tilbøjelige til luftvejs- og bihuleinfektioner.
En anden sygdom relateret til unormal udvikling af axoneme er polycystisk nyresygdom. I dette udvikles flere cyster i nyrerne, der ender med at ødelægge nyrerne. Denne sygdom skyldes en mutation i de gener, der koder for proteiner kaldet polycystiner.


Endnu ingen kommentarer