Det ketogenese Det er den proces, hvorved acetoacetat, β-hydroxybutyrat og acetone opnås, som sammen kaldes ketonlegemer. Denne komplekse og finregulerede mekanisme finder sted i mitokondrier, fra katabolisme af fedtsyrer.
At få ketonlegemer finder sted, når kroppen udsættes for udtømmende fasteperioder. Selvom disse metabolitter for det meste syntetiseres i leverceller, findes de som en vigtig energikilde i forskellige væv, såsom skeletmuskulatur og i hjerte- og hjernevæv..
Β-Hydroxybutyrat og acetoacetat er metabolitter, der anvendes som substrater i hjertemusklen og hjernebarken. I hjernen bliver ketonlegemer vigtige energikilder, når kroppen har tømt sit glukoselager.
Artikelindeks
Ketogenese betragtes som en meget vigtig fysiologisk funktion eller metabolisk vej. Generelt finder denne mekanisme sted i leveren, skønt det har vist sig, at den kan udføres i andre væv, der er i stand til at metabolisere fedtsyrer.
Dannelsen af ketonlegemer er den vigtigste metaboliske afledning af acetyl-CoA. Denne metabolit opnås fra den metaboliske vej kendt som β-oxidation, hvilket er nedbrydningen af fedtsyrer.
Tilgængeligheden af glucose i vævene, hvor β-oxidation opstår, bestemmer den metaboliske skæbne for acetyl-CoA. I særlige situationer er de oxiderede fedtsyrer næsten udelukkende rettet mod syntese af ketonlegemer..
Det vigtigste ketonlegeme er acetoacetat eller aceteddikesyre, som hovedsageligt syntetiseres i leverceller. De andre molekyler, der udgør ketonlegemerne, stammer fra acetoacetat.
Reduktionen af aceteddikesyre giver anledning til D-β-hydroxybutyrat, det andet ketonlegeme. Aceton er en forbindelse, der er vanskelig at nedbryde og produceres ved en spontan decarboxyleringsreaktion af acetoacetat (så det ikke kræver indgriben af et enzym), når det er til stede i høje koncentrationer i blodet.
Betegnelsen af ketonlegemer er blevet fastlagt ved konvention, da β-hydroxybutyrat strengt taget ikke har en ketonfunktion. Disse tre molekyler er opløselige i vand, hvilket letter deres transport i blodet. Dets vigtigste funktion er at give energi til visse væv, såsom skelet- og hjertemuskel.
Enzymerne, der er involveret i dannelsen af ketonlegemer, findes hovedsageligt i lever- og nyreceller, hvilket forklarer, hvorfor disse to placeringer er de største producenter af disse metabolitter. Dens syntese forekommer udelukkende og udelukkende i den mitokondrie matrix af celler.
Når disse molekyler er syntetiseret, passerer de ind i blodbanen og går til vævet, der kræver dem, hvor de nedbrydes til acetyl-CoA..
Den metaboliske skæbne for acetyl-CoA fra β-oxidation afhænger af kroppens metaboliske krav. Dette oxideres til COto og HtoEller via citronsyrecyklussen eller syntesen af fedtsyrer, hvis metabolismen af lipider og kulhydrater er stabil i kroppen.
Når kroppen har brug for dannelsen af kulhydrater, anvendes oxaloacetat til fremstilling af glukose (gluconeogenese) i stedet for at starte citronsyrecyklussen. Dette sker, som nævnt, når kroppen har en vis manglende evne til at få glukose, i tilfælde som langvarig faste eller tilstedeværelsen af diabetes.
På grund af dette anvendes acetyl-CoA som følge af oxidation af fedtsyrer til produktion af ketonlegemer..
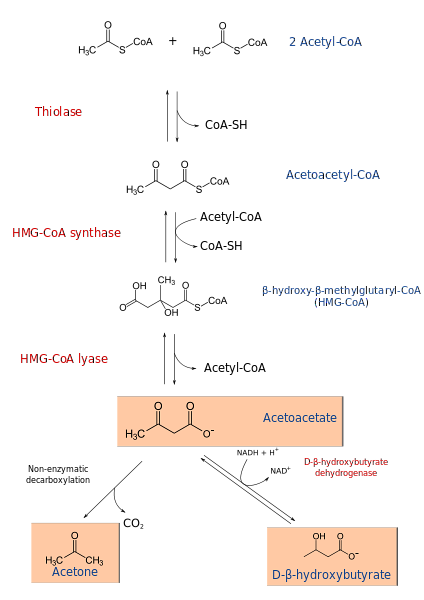
Ketogeneseprocessen begynder fra produkterne af β-oxidation: acetacetyl-CoA eller acetyl-CoA. Når substratet er acetyl-CoA, består det første trin af kondensering af to molekyler, en reaktion katalyseret af acetyl-CoA transferase, til dannelse af acetacetyl-CoA.
Acetacetyl-CoA kondenseres med en tredje acetyl-CoA gennem virkningen af HMG-CoA-syntase til dannelse af HMG-CoA (β-hydroxy-β-methylglutaryl-CoA). HMG-CoA nedbrydes til acetoacetat og acetyl-CoA ved virkningen af HMG-CoA lyase. På denne måde opnås det første ketonlegeme.
Acetoacetat reduceres til β-hydroxybutyrat ved intervention af β-hydroxybutyratdehydrogenase. Denne reaktion afhænger af NADH.
Det største acetoacetatketonlegeme er en β-ketosyre, som gennemgår ikke-enzymatisk decarboxylering. Denne proces er enkel og producerer acetone og COto.
Denne række reaktioner giver således anledning til ketonlegemer. Disse er opløselige i vand kan let transporteres gennem blodcirkulationen uden behov for at blive forankret til en albuminstruktur, som det er tilfældet med fedtsyrer, der er uopløselige i vandigt medium..
Fedtsyremetabolisme producerer substraterne for ketogenese, så disse to veje er funktionelt relaterede.
Acetoacetyl-CoA er en hæmmer af fedtsyremetabolisme, da det stopper aktiviteten af acyl-CoA dehydrogenase, som er det første enzym af β-oxidation. Derudover udøver den også hæmning af acetyl-CoA transferase og HMG-CoA syntase.
Enzymet HMG-CoA-syntase, underordnet af CPT-I (et enzym involveret i produktionen af acylcarnitin i β-oxidation), spiller en vigtig regulatorisk rolle i dannelsen af fedtsyrer.
Fodring af organismer regulerer et komplekst sæt hormonelle signaler. Kulhydrater, aminosyrer og lipider, der indtages i kosten, deponeres i form af triacylglyceroler i fedtvæv. Insulin, et anabolt hormon, er involveret i syntesen af lipider og dannelsen af triacylglyceroler.
På mitokondrie niveau styres β-oxidation ved indgang og deltagelse af nogle substrater i mitokondrierne. CPT I-enzymet syntetiserer Acyl Carnitin fra cytosolisk Acyl CoA.
Når kroppen fodres, aktiveres Acetyl-CoA-carboxylase, og citrat øger CPT I-niveauer, mens dets phosphorylering falder (reaktion afhængig af cyklisk AMP).
Dette forårsager en ophobning af malonyl CoA, som stimulerer syntesen af fedtsyrer og blokerer deres oxidation, hvilket forhindrer en forgæves cyklus i at blive genereret..
I tilfælde af faste er carboxylase-aktiviteten meget lav, da niveauerne af CPT I-enzymet er blevet reduceret, og det er også blevet phosphoryleret, hvilket aktiverer og fremmer oxidationen af lipider, hvilket efterfølgende vil tillade dannelse af ketonlegemer gennem acetyl- CoA.
Ketonlegemer diffunderer ud af cellerne, hvor de blev syntetiseret og transporteres til perifert væv i blodbanen. I disse væv kan de oxideres gennem tricarboxylsyrecyklussen.
I perifert væv oxideres β-hydroxybutyrat til acetoacetat. Derefter aktiveres det tilstedeværende acetoacetat ved virkningen af enzymet 3-ketoacyl-CoA transferase.
Succinyl-CoA fungerer som en CoA-donor ved at konvertere sig til succinat. Aktivering af acetoacetat forekommer for at forhindre succinyl-CoA i at omdanne til succinat i citronsyrecyklussen med den koblede syntese af GTP ved virkningen af succinyl-CoA-syntase.
Den resulterende acetoacetyl-CoA gennemgår en thiolytisk nedbrydning og producerer to acetyl-CoA-molekyler, der er inkorporeret i tricarboxylsyrecyklussen, bedre kendt som Krebs-cyklussen..
Leverceller mangler 3-ketoacyl-CoA transferase, hvilket forhindrer denne metabolit i at blive aktiveret i disse celler. På denne måde garanteres det, at ketonlegeme ikke oxiderer i cellerne, hvor de blev produceret, men at de kan overføres til vævene, hvor deres aktivitet er påkrævet..
I den menneskelige krop kan høje koncentrationer af ketonlegemer i blodet forårsage specielle tilstande kaldet acidose og ketonæmi..
Fremstillingen af disse metabolitter svarer til katabolismen af fedtsyrer og kulhydrater. En af de mest almindelige årsager til en patologisk ketogenese-tilstand er den høje koncentration af eddikesyre-dicarbonatfragmenter, der ikke nedbrydes af oxidationsvejen for tricarboxylsyrer..
Som en konsekvens er der en stigning i niveauerne af ketonlegemer i blodet over 2 til 4 mg / 100 N og deres tilstedeværelse i urinen. Dette resulterer i forstyrrelse af den mellemliggende metabolisme af disse metabolitter..
Visse defekter i hypofysen neuroglandulære faktorer, der regulerer nedbrydning og syntese af ketonlegemer sammen med lidelser i metabolismen af kulbrinter, er årsagen til tilstanden af hyperketonæmi..
Diabetes mellitus (type 1) er en endokrin sygdom, der forårsager øget produktion af ketonlegemer. Utilstrækkelig insulinproduktion deaktiverer transport af glukose til muskler, lever og fedtvæv og akkumuleres således i blodet.
Celler i fravær af glukose begynder processen med glukoneogenese og nedbrydning af fedt og protein for at genoprette deres stofskifte. Som en konsekvens falder oxaloacetatkoncentrationerne, og lipidoxidationen øges..
En akkumulering af acetyl-CoA opstår derefter, som i fravær af oxaloacetat ikke kan følge citronsyrevejen, hvilket forårsager den høje produktion af ketonlegemer, der er karakteristisk for denne sygdom..
Akkumuleringen af acetone detekteres ved dets tilstedeværelse i urinen og ånde hos mennesker med denne tilstand og er faktisk et af symptomerne, der indikerer manifestationen af denne sygdom.


Endnu ingen kommentarer