Det phosphatidylethanolamin (PE) er et glycerophospholipid, der er rigeligt i plasmamembranerne i prokaryote organismer. Tværtimod er det i eukaryote cellemembraner det næstmest forekommende glycerophospholipid på indersiden af plasmamembranen efter phosphatidylcholin..
På trods af overflod af phosphatidylethanolamin afhænger dens overflod ikke kun af celletypen, men også af rummet og tidspunktet for den specifikke cellelevecyklus, der overvejes..
Biologiske membraner er barrierer, der definerer cellulære organismer. Ikke kun har de beskyttelses- og isolationsfunktioner, men de er også nøglen til etablering af proteiner, der kræver et hydrofobt miljø for at de fungerer optimalt..
Både eukaryoter og prokaryoter har membraner, der hovedsageligt består af glycerophospholipider og i mindre grad sfingolipider og steroler..
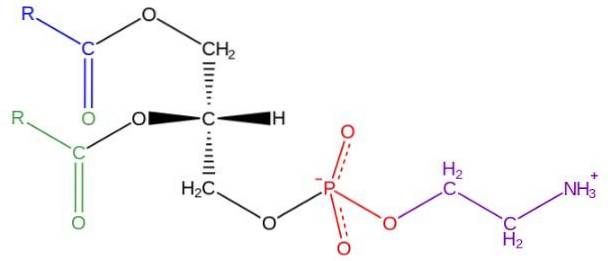
Glycerophospholipider er amfipatiske molekyler struktureret på en L-glycerol-rygrad, der esterificeres i sn-1 og sn-2-positionerne med to fedtsyrer med varierende længde og grad af mætning. I hydroxylen i sn-3-positionen esterificeres den med en phosphatgruppe, hvortil der igen kan knyttes forskellige typer molekyler, der giver anledning til de forskellige klasser af glycerophospholipider.
I den cellulære verden er der en lang række glycerophospholipider, men de mest rigelige er phosphatidylcholin (PC), phosphatidylethanolamin (PE), phosphatidylserin (PS), phosphatidylinositol (PI), phosphatidic acid (PA), phosphatidylglycerol (PG) og cardiolipin (CL).
Artikelindeks
Strukturen af phosphatidylethanolamin blev opdaget af Baer et al. I 1952. Som det er blevet eksperimentelt bestemt for alle glycerophospholipider, dannes phosphatidylethanolamin af et glycerolmolekyle forestret ved sn-1 og sn-2 positioner med syrekæder fede med mellem 16 og 20 kulstofatomer.
Fedtsyrerne forestret i sn-1 hydroxyl er generelt mættede (uden dobbeltbindinger) med maksimale længder på 18 carbonatomer, mens kæderne forbundet i sn-2-positionen er længere og med en eller flere umættelser (dobbeltbindinger).
Graden af mætning af disse kæder bidrager til membranens elasticitet, som har stor indflydelse på indsættelse og sekvestrering af proteiner i dobbeltlaget..
Phosphatidylethanolamin betragtes som et ikke-lamellært glycerophospholipid, da det har en konisk geometrisk form. Denne form er givet af den lille størrelse af dens polære gruppe eller "hoved" i forhold til den af fedtsyrekæderne, der omfatter de hydrofobe "haler".
"Hovedet" eller den polære gruppe af phosphatidylethanolamin har en zwitterionisk karakter, det vil sige den har grupper, der kan være positivt og negativt ladet under visse pH-betingelser..
Denne egenskab gør det muligt at hydrogenbinde med et stort antal aminosyrerester, og dens ladningsfordeling er en væsentlig determinant for domænetopologien for mange integrerede membranproteiner..
I eukaryote celler er syntesen af strukturelle lipider geografisk begrænset, idet det vigtigste biosyntese-sted er det endoplasmatiske retikulum (ER) og i mindre grad Golgi-apparatet..
Der er fire uafhængige biosyntetiske veje til produktion af phosphatidylethanolamin: (1) CDP-ethanolaminvejen, også kendt som Kennedy-vejen; (2) PSD-banen til phosphatidylserin (PS) decarboxylering; (3) acylering af lyso-PE og (4) baseændringsreaktioner i den polære gruppe af andre glycerophospholipider.
Phosphatidylethanolamin-biosyntese ved denne vej er begrænset til ER, og det er blevet vist, at det i hamsterleverceller er den vigtigste produktionsvej. Den består af tre på hinanden følgende enzymatiske trin katalyseret af tre forskellige enzymer.
I det første trin produceres phosphoethanolamin og ADP takket være virkningen af ethanolamin-kinase, som katalyserer den ATP-afhængige phosphorylering af ethanolamin..
I modsætning til planter er hverken pattedyr eller gær i stand til at producere dette substrat, så det skal indtages i kosten eller opnås ved nedbrydning af allerede eksisterende phosphatidylethanolamin- eller sfingosinmolekyler..
Phosphoethanolamin anvendes af CTP: phosphoethanolamin cytidyltransferase (ET) til dannelse af højenergiforbindelsen CDP: ethanolamin og et uorganisk phosphat.
1,2-Diacylglycerolethanolaminphosphotransferase (ETP) bruger energien indeholdt i CDP-ethanolaminbindingen til kovalent at binde ethanolamin til et membranindsat diacylglycerolmolekyle, hvilket giver anledning til phosphatidylethanolamin..
Denne rute fungerer både i prokaryoter og i gær og pattedyr. Hos bakterier forekommer det i plasmamembranen, men i eukaryoter forekommer det i et område af det endoplasmatiske retikulum, der er tæt beslægtet med mitokondriemembranen.
I pattedyr katalyseres vejen af et enkelt enzym, phosphatidylserin-decarboxylase (PSD1p), som er indlejret i mitokondrie-membranen, hvis gen er kodet af kernen. Reaktionen involverer decarboxylering af PS til phosphatidylethanolamin.
De resterende to veje (PE-lyso-acylering og polar gruppeafhængig calciumudveksling) forekommer i det endoplasmatiske retikulum, men bidrager ikke signifikant til den totale produktion af phosphatidylethanolamin i eukaryote celler..
Glycerophospholipider har tre hovedfunktioner i cellen, blandt hvilke strukturelle funktioner, energilagring og cellesignalering skiller sig ud..
Phosphatidylethanolamin er forbundet med forankring, stabilisering og foldning af flere membranproteiner samt de konformationsændringer, der er nødvendige for, at mange enzymer fungerer.
Der er eksperimentelle beviser, der foreslår phosphatidylethanolamin som et afgørende glycerophospholipid i det sene stadium af telophase under dannelsen af den kontraktile ring og etableringen af fragmoplast, der muliggør opdeling af membranen af de to datterceller..
Det har også en vigtig rolle i alle processer med fusion og fission (forening og adskillelse) af membranerne i både det endoplasmatiske retikulum og Golgi-apparatet..
I E. coli er det blevet vist, at phosphatidylethanolamin er nødvendig for den korrekte foldning og funktion af enzymet lactosepermease, hvorfor det er blevet foreslået, at det spiller en rolle som en molekylær "chaperone".
Phosphatidylethanolamin er den vigtigste donor til ethanolaminmolekylet, der er nødvendigt til posttranslationel modifikation af adskillige proteiner, såsom GPI-ankre.
Dette glycerophospholipid er forløberen for adskillige molekyler med enzymatisk aktivitet. Derudover kan molekyler afledt af dets metabolisme såvel som diacylglycerol, phosphatidsyre og nogle fedtsyrer fungere som anden budbringere. Derudover er det et vigtigt substrat til produktionen af phosphatidylcholin..

Endnu ingen kommentarer