Det isogami det er et reproduktionssystem af planter, hvor kønscellerne er morfologisk ens. Lighed opstår i form og størrelse, og manlige og kvindelige kønsceller kan ikke skelnes. Dette reproduktive system betragtes som forfædres. Det forekommer i forskellige grupper af alger, svampe og protozoer.
De gameter, der er involveret i isogami, kan være mobile (cilierede) eller ej. Deres forening finder sted ved bøjning. Udifferentierede kønsceller smelter sammen og udveksler genetisk materiale.
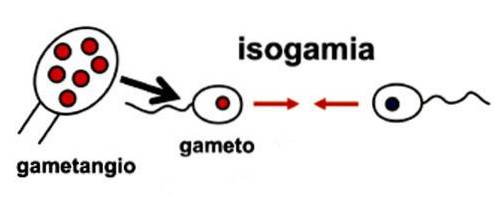
Isogami kan være homotisk eller heterotisk. Det er homotisk, når fusionen opstår mellem kønsceller, der har samme genom. I heterotisk isogami har kønsceller en anden genetisk sammensætning.
Artikelindeks
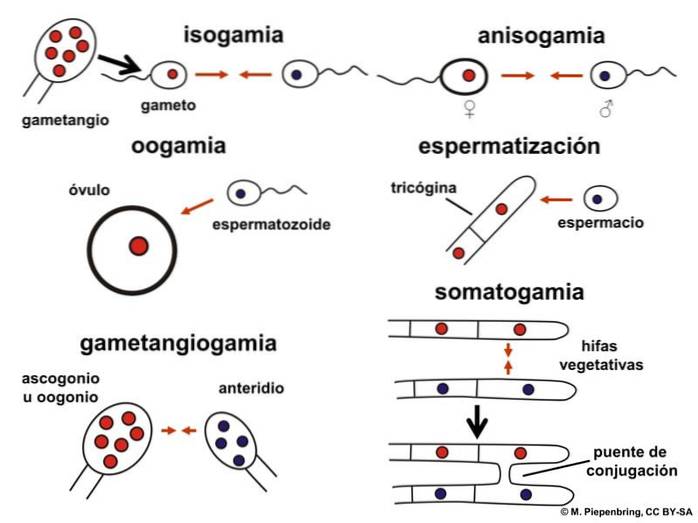
Reproduktion ved isogami sker ved konjugation. I dette bevæger indholdet af en celle sig mod en anden, og der opstår fusion.
Processer med carigami (fusion af kerner) og plasmogami (fusion af cytoplasma) er involveret. Somatisk celledifferentiering i seksuelle celler kan være forbundet med miljømæssige forhold. Interaktion med andre individer af samme art kan også påvirke.
Efter differentiering opstår, skal kønscellerne finde og genkende andre kønsceller. I grupper, hvor isogami forekommer, finder genkendelse og fusion af kønsceller sted på forskellige måder.
Sexceller kan flagelleres eller immobile. I nogle tilfælde er de store, som i nogle grønalger.
Der er to typer isogami relateret til den genetiske sammensætning af kønsceller.
En persons gamet er konjugeret med en anden fra den samme klongruppe. I dette tilfælde anses selvbefrugtning for at forekomme.
Alle kerner har den samme genotype, og der er ingen interaktion med en anden genotype. Somatiske celler differentieres direkte i kønsceller.
Kønsceller dannes i klonpopulationer, og senere forekommer fusion for at danne zygoten.
Kønsceller produceres i forskellige individer, som har en anden genetisk sammensætning.
Det kræves, at kønsceller har genetisk kompatibilitet, for at fusion kan forekomme. To typer gameter dannes generelt. "Plus" og "minus", der er kompatible med hinanden.
Den gametangiale celle (som producerer gameten) af den ene type danner et par med den for den anden type. Disse genkendes gennem kemisk kommunikation, der i nogle tilfælde involverer feromonproduktion.
Betingelsen for isogami ser ud til at dominere i encellede organismer, mens anisogami næsten er universel for multicellulære eukaryoter. I de fleste eukaryote slægter af encellede organismer er kønscellerne lige store, og vi skelner ikke mellem mænd og kvinder..
I eukaryoter er der et betydeligt antal arter med isogamiske kønsceller. Imidlertid vil vi kun nævne de slægter, der konstant vises i den biologiske litteratur - selvom der er mange flere..
Den velkendte sociale amøbe af arten Dictyostelium discoideum, den almindelige gær, vi bruger til at lave mad Saccharomyces cerevisiae og den protozoiske parasit, der forårsager søvnsygdom Trypanosoma brucei er alle eksempler på organismer med identiske kønsceller.
I grønalger er isogami et almindeligt fænomen. Faktisk er der to typer isogami i disse organismer.
Nogle arter producerer relativt mellemstore kønsceller med et fototaktisk system repræsenteret af en øjenplet. Andre arter har de samme kønsceller, men meget mindre end i det foregående tilfælde. De mangler også øjenpletten.
Det er imidlertid ikke muligt at foretage en sådan radikal observation og begrænse isogamiske kønsceller til encellede slægter og anisogamic til flercellede væsener..
Faktisk præsenterer planter nogle undtagelser fra denne regel, da koloniale grønne alger, såsom Pandorina, Volvulina Y Yamagishiella præsentere betingelsen for isogami.
Der er også undtagelser i den modsatte retning, da der er encellede organismer, såsom de grønne alger af ordenen Bryopsidales, der præsenterer forskellige kønsceller..
I alger er tilstedeværelsen af to typer kønsceller forbundet med isogami blevet observeret.
I nogle grupper er kønscellerne mellemstore og har fototaksmekanismer. Der er en øjenplet, der stimuleres af lys.
De er generelt forbundet med tilstedeværelsen af kloroplaster og evnen til at akkumulere reserve stoffer. I andre tilfælde er kønscellerne meget små og har ikke et øje..
Seksuel reproduktion i isogami alger forekommer forskelligt.
Det er en gruppe af encellede grønalger med to flageller. Det præsenterer heterotisk isogami. Homotisk isogami kan forekomme i nogle arter.
Haploide vegetative celler differentierer til kønsceller, når nitrogenforholdene stiger i mediet. Der er to typer gameter med forskellige genetiske komplement.
Kønsceller producerer agglutininer (adhæsionsmolekyler), der fremmer bindingen af flageller. Efter fusion giver de to gameter den genetiske information, der er nødvendig for embryonets udvikling.
Disse alger hører til Charyophyta-divisionen. De er encellede. De præsenterer homotisk og heterotisk isogami.
Gameter er ikke mobile. I dette tilfælde, når kønscellerne stammer, dannes en bøjningspapille. Cytoplasmer frigives ved brud på cellevæggen.
Derefter forekommer fusionen af protoplasmerne i begge kønsarter, og zygoten dannes. Kemisk tiltrækning mellem de forskellige genetiske typer anses for at forekomme i heterotisk isogami.
De er flercellede organismer med flagellat-isogame kønsceller. Andre grupper reproducerer ved anisogami eller oogamy.
Kønsceller er morfologisk de samme, men de opfører sig anderledes. Der er arter, hvor den kvindelige type frigiver feromoner, der tiltrækker den mandlige type.
I andre tilfælde bevæger sig en type gamete i en kort periode. Indtag derefter flagellen og slip feromoner. Den anden type bevæger sig i længere tid og har en feromon signalreceptor.
Både homotiske og heterotiske typer er isogami. I de fleste tilfælde er kønsgenkendelse forbundet med produktionen af feromoner.
I forskellige encellede grupper såsom Saccharomyces, gameter differentieres som reaktion på en ændring i sammensætningen af dyrkningsmediet. Somatiske celler deler sig ved meiose under visse betingelser, såsom lave kvælstofniveauer..
Kønsceller med forskellig genetisk sammensætning genkendes af feromonsignaler. Cellerne danner fremspring mod feromonkilden og slutter sig til deres apices. Kerne af begge gameter vandrer, indtil de smelter sammen og danner en diploid celle (zygote).
De er flercellede organismer. De præsenterer hovedsageligt heterotiske systemer. Under seksuel udvikling danner de donor- (mandlige) og modtagelige (kvindelige) strukturer.
Cellefusion kan forekomme mellem en hyfa og en mere specialiseret celle eller mellem to hyfer. Indtastningen af donorkernen (han) i hyfaen stimulerer udviklingen af en frugtkrop.
Kerner smelter ikke straks. Frugterlegemet danner en dikaryot struktur med kerner med forskellig genetisk sammensætning. Derefter smelter kernerne sammen og deler sig ved meiose.
Isogamy forekommer i flagellate encellede grupper. Disse cilierede organismer etablerer cytoplasmisk forbindelse mellem kønsceller i specialiserede områder af plasmamembranen..
De cilierede grupper har to kerner, en makronukleus og en mikronukleus. Makronukleus er den somatiske form. Den diploide mikronukleus deler sig ved meiose og danner gameten.
De haploide kerner udveksles af en cytoplasmatisk bro. Derefter gendannes cytoplasmaerne i hver celle, og de genvinder deres autonomi. Denne proces er unik inden for eukaryoter.
På Euplots Specifikke feromoner af hver genetisk type produceres. Celler stopper somatisk vækst, når de opdager et feromon med forskellig genetisk sammensætning.
For arter af Dileptus genkendelsesmolekyler præsenteres på celleoverfladen. Kompatible gameter er bundet af adhæsionsproteiner i cilier.
På Paramecium genkendelsesstoffer produceres mellem kompatible kønsceller. Disse stoffer fremmer foreningen af kønsceller såvel som deres adhæsion og efterfølgende fusion..
I evolutionær biologi er forældrenes investering et af de mest diskuterede emner, når vi taler om komplekse organismer (såsom pattedyr). Dette koncept blev udviklet af den fremtrædende biolog Sir Ronald Fisher i sin bog "Den genetiske teori om naturlig udvælgelse”, Og indebærer forældrenes udgifter til afkomets velfærd.
Ligestilling i kønsceller indebærer, at forældrenes investering vil være symmetrisk for begge organismer, der er involveret i den reproduktive begivenhed..
I modsætning til anisogami-systemet, hvor forældrenes investeringer er asymmetriske, og det er den kvindelige gamet, der tilvejebringer det meste af de ikke-genetiske ressourcer (næringsstoffer osv.) Til udvikling af zygote. Med udviklingen af systemer, der præsenterer dimorfisme i deres kønsceller, udviklede en asymmetri sig også i forældrenes organismer.
Ifølge beviser og reproduktionsmønstre, som vi finder hos moderne arter, synes det logisk at betragte isogami som den forfædres tilstand, der vises i de første faser af seksuel reproduktion..
I flere slægter af flercellede organismer, såsom planter og dyr, har et differentielt reproduktionssystem uafhængigt udviklet sig, hvor de kvindelige kønsceller er store og immobile, og de mandlige er små og med evnen til at bevæge sig til ægløsning..
Selvom de nøjagtige baner for ændring fra en isogamisk til en anisogamisk tilstand ikke er kendt, er flere teorier blevet formuleret..
En af dem fremhæver en mulig afvejning mellem størrelsen på kønscellerne og deres antal. Ifølge dette argument er anisogamiens oprindelse en evolutionært stabil strategi forårsaget af forstyrrende udvælgelse i søgen efter zygotens effektivitet og overlevelse..
En anden teori søger at forklare fænomenet som en måde at kompensere for en immobil celle (ægget) med mange celler med evnen til at bevæge sig (sædcellerne)..
En tredje opfattelse forklarer dannelsen af anisogami som en adaptiv egenskab for at undgå konflikter mellem kernen og cytoplasmaet på grund af den uniparental arv af organellerne..



Endnu ingen kommentarer