Det mitose det er en proces med celledeling, hvor en celle producerer genetisk identiske datterceller; for hver celle dannes to "døtre" med den samme kromosombelastning. Denne opdeling finder sted i de somatiske celler i eukaryote organismer..
Denne proces er et af stadierne i eukaryote organismernes cellecyklus, der består af 4 faser: S (DNA-syntese), M (celledeling), G1 og G2 (mellemfaser, hvor mRNA'er og proteiner produceres). Sammen betragtes G1-, G2- og S-faserne som en grænseflade. Nuklear og cytoplasmatisk opdeling (mitose og cytokinese) udgør den sidste fase af cellecyklussen.
På molekylært niveau initieres mitose ved aktivering af en kinase (protein) kaldet MPF (Maturation Promoting Factor) og den deraf følgende phosphorylering af et betydeligt antal af cellens komponentproteiner. Sidstnævnte tillader cellen at præsentere de morfologiske ændringer, der er nødvendige for at udføre delingsprocessen..
Mitose er en aseksuel proces, da stamcellen og dens døtre har nøjagtig den samme genetiske information. Disse celler er kendt som diploide, fordi de bærer den komplette kromosombelastning (2n).
Meiose er derimod processen med celledeling, der fører til seksuel reproduktion. I denne proces replikerer en diploid stamcelle sine kromosomer og deler sig derefter to gange i træk (uden at replikere dens genetiske information). Endelig genereres 4 datterceller med kun halvdelen af den kromosomale belastning, der kaldes haploide (n).
Artikelindeks
Mitose i encellede organismer producerer generelt datterceller, der meget ligner deres forfædre. I modsætning hertil kan denne proces under udviklingen af flercellede væsner give anledning til to celler med nogle forskellige egenskaber (på trods af at de er genetisk identiske)..
Denne celledifferentiering giver anledning til de forskellige celletyper, der udgør flercellede organismer..
I løbet af en organisms liv foregår cellecyklussen kontinuerligt og danner konstant nye celler, som igen vokser og forbereder sig på at dele sig gennem mitose.
Cellevækst og opdeling reguleres af mekanismer, såsom apoptose (programmeret celledød), som gør det muligt at opretholde en balance og undgår overskydende vævsvækst. På denne måde sikres det, at de defekte celler udskiftes med nye celler i henhold til kroppens krav og behov..
Evnen til at reproducere er en af de vigtigste egenskaber ved alle organismer (fra encellede til multicellulære) og cellerne, der komponerer den. Denne kvalitet sikrer kontinuiteten i dine genetiske oplysninger..
At forstå processerne med mitose og meiose har spillet en grundlæggende rolle i forståelsen af de spændende cellulære egenskaber ved organismer. F.eks. Egenskaben ved at holde antallet af kromosomer konstant fra en celle til en anden inden for et individ og mellem individer af samme art.
Når vi lider af en eller anden form for snit eller sår på vores hud, observerer vi, hvordan den beskadigede hud kommer sig i løbet af få dage. Dette sker takket være processen med mitose.
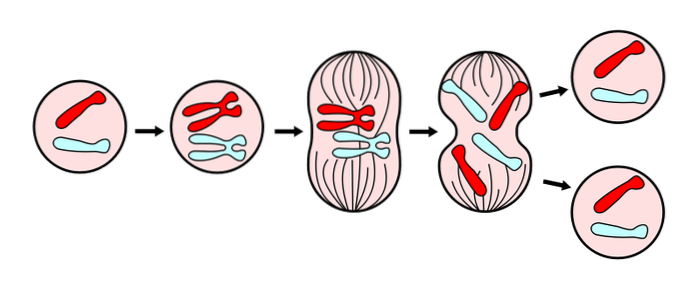
Generelt følger mitose den samme sekvens af processer (faser) i alle eukaryote celler. I disse faser forekommer mange morfologiske ændringer i cellen. Blandt dem kondens af kromosomer, brud på kernemembranen, adskillelse af cellen fra den ekstracellulære matrix og andre celler og opdeling af cytoplasmaet..
I nogle tilfælde betragtes nuklear opdeling og cytoplasmatisk opdeling som forskellige faser (henholdsvis mitose og cytokinese).
For en bedre undersøgelse og forståelse af processen er der udpeget seks (6) faser, kaldet: profase, prometaphase, metaphase, anaphase og telophase, idet cytokinesis betragtes som en sjette fase, som begynder at udvikle sig under anaphase.
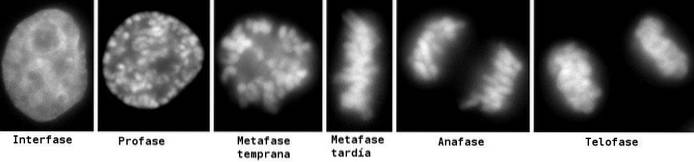
Disse faser er blevet undersøgt siden det 19. århundrede gennem lysmikroskopet, så i dag er de let genkendelige i henhold til de morfologiske egenskaber, som cellen præsenterer, såsom kromosomal kondensation og dannelsen af den mitotiske spindel..
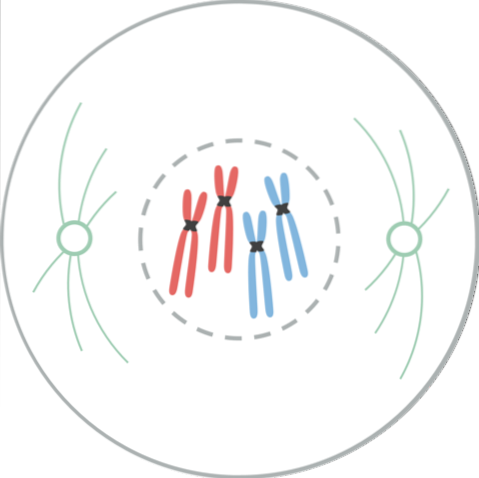
Profase er den første synlige manifestation af celledeling. I denne fase kan udseendet af kromosomer ses som skelneformer på grund af den gradvise komprimering af kromatin. Denne kondensation af kromosomer begynder med phosphorylering af Histone H1-molekyler med MPF-kinase..
Kondensationsprocessen består af sammentrækning og derfor reduktion af kromosomernes størrelse. Dette sker på grund af krøllefibrene, der producerer lettere forskydelige strukturer (mitotiske kromosomer).
Kromosomer, der tidligere blev duplikeret i S-perioden af cellecyklussen, får et dobbeltstrenget udseende, kaldet søsterkromatider, disse tråde holdes sammen gennem en region kaldet centromeren. I denne fase forsvinder nukleoli også.
Under profase dannes den mitotiske spindel, der består af mikrotubuli og proteiner, der udgør et sæt fibre.
Når spindlen dannes, adskilles mikrotubuli i cytoskeletet (ved at deaktivere proteinerne, der opretholder deres struktur), hvilket tilvejebringer det nødvendige materiale til dannelsen af den mitotiske spindel..
Centrosomet (en membranløs organel, funktionel i cellecyklussen), duplikeret ved grænsefladen, fungerer som samleenheden for spindelmikrotubuli. I dyreceller har centrosomet et par centrioler i midten; men disse er fraværende i de fleste planteceller.
De duplikerede centrosomer begynder at adskille sig fra hinanden, mens mikrotubuli i spindlen samles i hver af dem og begynder at migrere mod modsatte ender af cellen..
I slutningen af profasen begynder bruddet på den nukleare hylster, der forekommer i separate processer: adskillelse af nukleare porer, nukleare lameller og nukleare membraner. Denne pause gør det muligt for den mitotiske spindel og kromosomer at begynde at interagere..
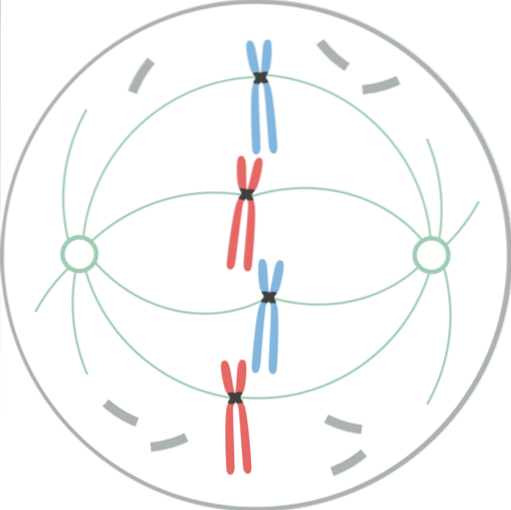
På dette stadium er kernekapslen blevet helt fragmenteret, så spindelens mikrotubuli invaderer dette område og interagerer med kromosomerne. De to centrosomer er adskilt, hver placeret ved polerne i den mitotiske spindel, i modsatte ender af cellerne..
Nu omfatter den mitotiske spindel mikrotubuli (som strækker sig fra hvert centrosom mod centrum af cellen), centrosomerne og et par asters (strukturer med en radial fordeling af korte mikrotubuli, der udfolder sig fra hvert centrosom)..
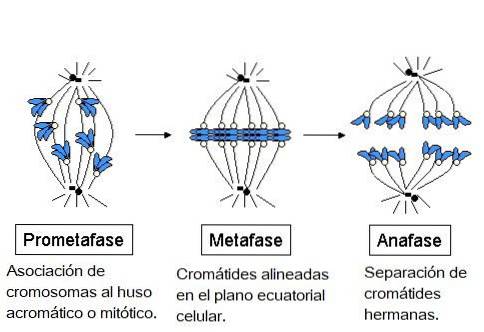
Kromatiderne udviklede hver især en specialiseret proteinstruktur, kaldet kinetochore, der ligger i centromeren. Disse kinetochore er placeret i modsatte retninger, og nogle mikrotubuli, kaldet mikrotubuli af kinetochore, overholder dem..
Disse mikrotubuli, fastgjort til kinetochore, begynder at bevæge sig til kromosomet fra den ende, hvorfra de strækker sig; nogle fra den ene pol og andre fra den modsatte pol. Dette skaber en "pull and shrink" -effekt, der, når den er stabiliseret, gør det muligt for kromosomet at ende op mellem enderne af cellen..
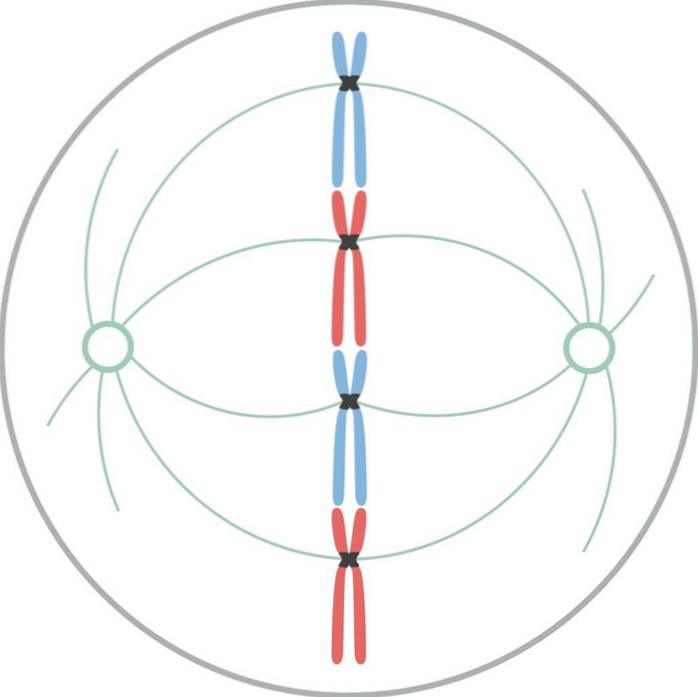
I metafase er centrosomer placeret i modsatte ender af celler. Spindlen viser en klar struktur, hvor kromosomerne er placeret i midten. Centromerer af disse kromosomer er bundet til fibrene og justeret i et imaginært plan kaldet metafasepladen..
Kromatidkinetochores forbliver knyttet til mikrotubuli i kinetochore. Mikrotubuli, der ikke klæber til kinetochores og strækker sig fra modsatte poler af spindlen, interagerer nu med hinanden. På dette tidspunkt er mikrotubuli fra asters i kontakt med plasmamembranen..
Denne vækst og interaktion mellem mikrotubuli fuldender strukturen i den mitotiske spindel og giver den et "fuglebur" -udseende..
Morfologisk er denne fase den med de mindste ændringer, hvorfor det blev betragtet som en hvilefase. Selvom de ikke er let synlige, forekommer der dog mange vigtige processer i det ud over at være det længste stadium af mitose.
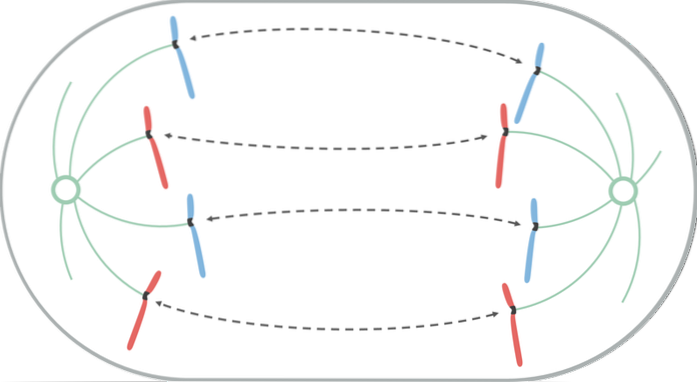
Under anafase begynder hvert par kromatider at adskille sig (på grund af inaktivering af proteinerne, der holder dem sammen). Separerede kromosomer bevæger sig til modsatte ender af cellen.
Denne migrationsbevægelse skyldes forkortelsen af kinetokorens mikrotubuli, hvilket genererer en "pull" -effekt, der får hvert kromosom til at bevæge sig fra dets centromer. Afhængigt af placeringen af centromeren på kromosomet kan det antage en bestemt V- eller J-form under dens forskydning..
Mikrotubuli, der ikke klæber sig til kinetochore, vokser og forlænges ved adhæsionen af tubulin (protein) og ved virkningen af motoriske proteiner, der bevæger sig over dem, så kontakten mellem dem stopper. Når de bevæger sig væk fra hinanden, gør polens spindler det også og forlænger cellen.
I slutningen af denne fase er grupperne af kromosomer placeret i modsatte ender af den mitotiske spindel, hvilket efterlader hver ende af cellen med et komplet og ækvivalent sæt kromosomer..
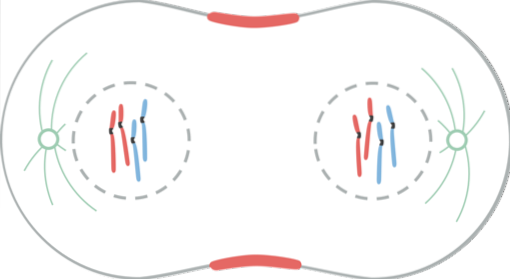
Telofasen er den sidste fase af nuklear division. Kinetochore mikrotubuli opløses, mens polære mikrotubuli forlænger sig længere.
Kernemembranen begynder at dannes omkring hvert sæt kromosomer ved hjælp af stamcellens nukleare hylstre, der var som vesikler i cytoplasmaet.
I dette trin dekondenseres de kromosomer, der er ved cellepolerne, grundet dephosphorylering af histonmolekylerne (H1). Dannelsen af kernerne i kernemembranen styres af flere mekanismer.
Under anafase begyndte mange af de phosphorylerede proteiner i profase at dephosphorylere. Dette gør det muligt i begyndelsen af telofasen, at de nukleare vesikler begynder at samles igen og forbinder sig med kromosomens overflade..
På den anden side samles den nukleare pore igen, hvilket tillader pumpning af nukleare proteiner. Proteinerne i det nukleare lag dephosphoryleres, hvilket tillader dem at associere igen for at afslutte dannelsen af det nukleare lamina.
Endelig, efter at kromosomerne er helt dekondenseret, genstartes RNA-syntese, hvorved dannes nucleoli igen og således fuldender dannelsen af de nye mellemfasekerner i dattercellerne..

Cytokinesis tages som en separat begivenhed fra nuklear opdeling, og almindeligvis i typiske celler ledsager den cytoplasmatiske delingsproces hver mitose, begyndende ved anafase. Flere undersøgelser har vist, at der i nogle embryoner forekommer flere nukleare opdelinger inden cytoplasmatisk opdeling.
Processen begynder med fremkomsten af en rille eller kløft, der er markeret i metafasepladens plan, hvilket sikrer, at opdelingen finder sted mellem kromosomgrupperne. Stedet for kløften er angivet med mitotisk spindel specifikt asterernes mikrotubuli.
I den markerede kløft findes en række mikrofilamenter, der danner en ring rettet mod den cytoplasmatiske side af cellemembranen, stort set sammensat af actin og myosin. Disse proteiner interagerer med hinanden, så ringen kan trække sig sammen omkring rillen..
Denne sammentrækning genereres ved at glide filamenterne af disse proteiner, når de interagerer med hinanden, på samme måde som de gør, for eksempel i muskelvæv..
Sammentrækningen af annulus uddybes og udøver en "klemme" -effekt, der endelig deler stamfadercellen og muliggør adskillelse af dattercellerne med deres udviklende cytoplasmiske indhold..
Planteceller har en cellevæg, så deres cytoplasmatiske delingsproces er forskellig fra den tidligere beskrevne og begynder i telofase..
Dannelsen af en ny cellevæg begynder, når mikrotubuli i den resterende spindel samles og udgør fragmoplasten. Denne cylindriske struktur består af to sæt mikrotubuli, der er forbundet i deres ender, og hvis positive poler er indlejret i et elektronisk kort i ækvatorialplanet..
Små vesikler fra Golgi-apparatet, pakket med cellevægsforløbere, bevæger sig gennem mikrotubuli i fragmoplasten til ækvatorialområdet og kombineres for at danne en celleplade. Indholdet af vesiklerne udskilles i denne plade, når den vokser.
Denne plak vokser og smelter sammen med plasmamembranen langs celleomkredsen. Dette sker på grund af den konstante omlejring af fragmoplastens mikrotubuli ved periferien af pladen, hvilket tillader flere vesikler at bevæge sig mod dette plan og tømme deres indhold..
På denne måde forekommer den cytoplasmatiske adskillelse af dattercellerne. Endelig tillader indholdet af cellepladen sammen med cellulosemikrofibrene i den at fuldføre dannelsen af den nye cellevæg..
Mitose er en mekanisme for opdeling i celler og er en del af en af faserne i cellecyklussen i eukaryoter. På en enkel måde kan vi sige, at den vigtigste funktion ved denne proces er reproduktion af en celle i to datterceller.
For encellede organismer betyder celledeling generering af nye individer, mens for multicellulære organismer er denne proces en del af væksten og korrekt funktion af hele organismen (celledeling genererer udvikling af væv og vedligeholdelse af strukturer).
Mitoseprocessen aktiveres i henhold til organismen. I pattedyr begynder for eksempel røde blodlegemer (erytrocytter) at dele sig og danner flere celler, når kroppen har brug for bedre iltoptagelse. Tilsvarende reproducerer hvide blodlegemer (leukocytter), når det er nødvendigt at bekæmpe en infektion..
I modsætning hertil mangler nogle specialiserede dyreceller praktisk taget processen med mitose, eller det er meget langsomt. Eksempler på dette er nerveceller og muskelceller).
Generelt er de celler, der er en del af kroppens bindevæv og strukturelle væv, og hvis reproduktion kun er nødvendig, når en celle har en defekt eller forringelse og skal udskiftes.
Celledelings- og vækstkontrolsystemet er meget mere komplekst i flercellede organismer end i encellede. I sidstnævnte er reproduktion grundlæggende begrænset af tilgængeligheden af ressourcer.
I dyreceller arresteres deling, indtil der er et positivt signal, der aktiverer denne proces. Denne aktivering kommer i form af kemiske signaler fra naboceller. Dette gør det muligt at forhindre ubegrænset vækst af væv og reproduktion af defekte celler, som alvorligt kan skade organismenes liv.
En af mekanismerne, der styrer cellemultiplikation, er apoptose, hvor en celle dør (på grund af produktionen af visse proteiner, der aktiverer selvdestruktion), hvis den udgør betydelig skade eller er inficeret af en virus.
Der er også regulering af celleudvikling gennem inhibering af vækstfaktorer (såsom proteiner). Således forbliver cellerne ved grænsefladen uden at gå videre til M-fasen i cellecyklussen..
Mitoseprocessen finder sted i langt størstedelen af eukaryote celler, fra encellede organismer såsom gær, der bruger den som en aseksuel reproduktionsproces, til komplekse flercellede organismer såsom planter og dyr..
Skønt cellecyklussen generelt er den samme for alle eukaryote celler, er der bemærkelsesværdige forskelle mellem encellede og multicellulære organismer. I den førstnævnte foretrækkes vækst og deling af celler ved naturlig selektion. I flercellede organismer er spredning begrænset af strenge kontrolmekanismer.
I encellede organismer forekommer reproduktion på en accelereret måde, da cellecyklussen fungerer konstant, og dattercellerne hurtigt går i retning mod mitose for at fortsætte med denne cyklus. Mens celler fra flercellede organismer tager betydeligt længere tid at vokse og dele sig.
Der er også nogle forskelle mellem de mitotiske processer af plante- og dyreceller, som i nogle af faserne i denne proces, men i princippet fungerer mekanismen på en lignende måde i disse organismer..
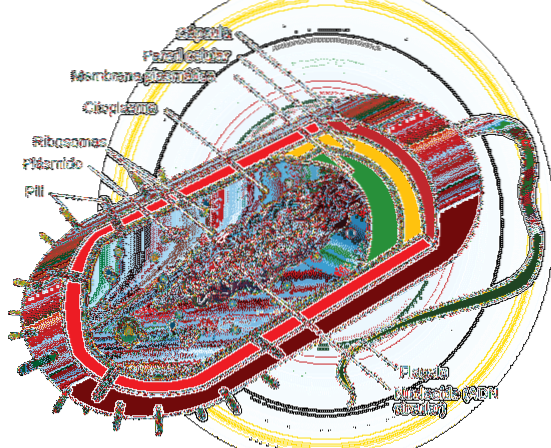
Prokaryote celler vokser og deler sig generelt hurtigere end eukaryote celler.
Organismer med prokaryote celler (generelt encellede eller i nogle tilfælde multicellulære) mangler en nuklear membran, der isolerer det genetiske materiale i en kerne, så det er spredt i cellen i et område kaldet nucleoid. Disse celler har et cirkulært hovedkromosom.
Celledeling i disse organismer er således meget mere direkte end i eukaryote celler, der mangler den beskrevne mekanisme (mitose). I dem udføres reproduktion ved en proces kaldet binær fission, hvor DNA-replikation begynder på et specifikt sted på det cirkulære kromosom (replikationsorigin eller OriC)..
To oprindelser dannes derefter, der migrerer til modsatte sider af cellen, når replikation opstår, og cellen strækker sig til dobbelt så stor. I slutningen af replikationen vokser cellemembranen ind i cytoplasmaet og deler stamcellen i to døtre med det samme genetiske materiale.
Udviklingen af eukaryote celler medførte en stigning i kompleksiteten i genomet. Dette involverede udviklingen af mere detaljerede delingsmekanismer.
Der er hypoteser, der foreslår, at bakteriedeling er forgængermekanismen for mitose. Der er fundet et vist forhold mellem proteiner associeret med binær fission (som kan være dem, der forankrer kromosomer til specifikke steder på plasmamembranen hos døtre) med tubulin og actin i eukaryote celler..
Nogle undersøgelser indikerer visse særegenheder i opdelingen af moderne encellede protister. I dem forbliver kernemembranen intakt under mitose. De replikerede kromosomer forbliver forankret på visse steder på denne membran og adskiller sig, når kernen begynder at strække sig under celledeling..
Dette viser noget sammenfald med processen med binær fission, hvor de replikerede kromosomer binder sig til bestemte steder på cellemembranen. Hypotesen foreslår derefter, at protisterne, der præsenterer denne kvalitet under deres celledeling, kunne have opretholdt denne egenskab ved en forfædren prokaryot celle..
På nuværende tidspunkt er der endnu ikke udviklet forklaringer på, hvorfor det i eukaryote celler fra flercellede organismer er nødvendigt for den nukleare membran at gå i opløsning under processen med celledeling..

Endnu ingen kommentarer