
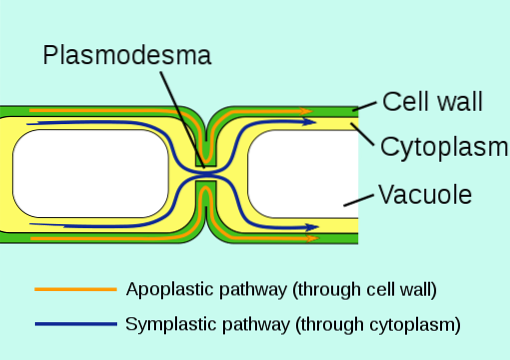
Det plasmodesmer er cytosoliske forbindelser, der opstår mellem tilstødende planteceller, det vil sige de kommunikerer protoplasterne (plasmamembran og cytosol) gennem cellevæggen og danner et forenklet kontinuum.
Disse strukturer er funktionelt analoge eller ækvivalente med kløftkryds (hulkryds), der observeres mellem cellerne i et dyrevæv, og deres vigtigste funktion er at kommunikere cellerne med hinanden og tjene som en kanal til transport af forskellige typer ioner og molekyler.
Plasmodesmata blev beskrevet for mere end 100 år siden af Tangl, og siden da er der blevet offentliggjort hundreder af undersøgelser, hvor deres funktionsmekanisme, deres struktur og andre relaterede aspekter er detaljeret detaljeret..
På nuværende tidspunkt er det kendt, at disse cytosoliske "kanaler" eller "forbindelser" mellem celler er strukturer under strenge kontrolmekanismer, og det er også blevet bestemt, at de hovedsageligt er sammensat af integrerede membranproteiner, chaperoneproteiner og andre proteiner specialiseret i transport af stoffer..
Artikelindeks
Plasmodesmata forbinder celler, der hører til det samme "forenklede domæne" i et plantevæv, hvilket betyder, at ikke alle celler i en plante er forbundet med hinanden, men der er forskellige specifikke "regioner" i et væv, hvor cellerne der findes der udveksler information permanent.
Disse er meget dynamiske strukturer; deres antal, struktur og funktion kan modificeres som reaktion på et specifikt funktionelt krav til et væv.
Derudover kan disse kanaler nedbrydes eller "forsegles" i nogle cellulære grænseflader (rummet mellem to celler), hvilket indebærer dannelse af en forenklet "barriere" mellem cellerne i nogle plantevæv og favoriserer isolering af definerede regioner i en væv.
Nogle bibliografiske citater antyder, at plasmodesmata er strukturer så komplekse som de såkaldte nukleare pore-komplekser, som udfører lignende funktioner, men i translokation af molekylær information fra det cytosoliske miljø til det indre af kernen..
Et hurtigt blik på et plantevæv er nok til at kontrollere, at der findes flere typer plasmodesmata..
Ifølge nogle forfattere kan disse klassificeres som primære og sekundære i henhold til det øjeblik de dannes i løbet af en celle; eller så enkelt og forgrenet, afhængigt af morfologien for de kanaler, der dannes mellem celle og celle.
Uanset hvilken type plasmodesmus det drejer sig om, er dens "strukturelle arkitektur" mere eller mindre ækvivalent, da det næsten altid er et spørgsmål om kanaler med en diameter, der varierer mellem 20 og 50 nm, hvis indgange eller åbninger er lidt mere snævre, udgør det, der er kendt som en "indsnævring af flaskehalsen".
Nogle forskere har foreslået, at en sådan indsnævring i plasmodesmata-åbningerne deltager i reguleringen af strømmen af stoffer gennem dem, det vil sige, at deres udvidelse (ekspansion) eller indsnævring (reduktion i diameter) bestemmer mængden og hastigheden af strømningen..
Disse "flaskehalse" er sammensat af et stof kendt som callose (β-1,3-glucan), og som det kan udledes, findes de i de områder, der er tættest på væggen af planteceller forbundet med disse kanaler..
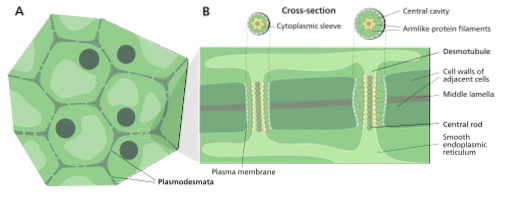
Primær plasmodesmata dannes i "cellepladen" under cytokinesis, hvilket er tidspunktet for mitose, hvor de to datterceller adskiller sig. Disse kan dog gennemgå strukturændringer og ændre deres distribution og drift under udviklingen af det anlæg, som de tilhører..
Disse plasmodesmata er faktisk membranmiljøer, der består af porer i plasmamembranen, der danner en slags bro mellem cellevæggen og et aksialt element i det "fangede" endoplasmatiske retikulum kendt som desmotubuli..
En demotubule er en cylindrisk struktur med mere eller mindre 15 nm i diameter, sammensat af det endoplasmatiske retikulum i en celle, der er kontinuerlig med cisternerne i det endoplasmatiske reticulum i den nærliggende celle, der er forbundet via plasmodesmet..
Mellem "streng" repræsenteret af demotubuli og plasmamembranen, der udgør det cylindriske hulrum, der er plasmodesmus, er der et rum kendt som "cytoplasmisk muffe" (fra engelsk Cytoplasmisk ærme), hvorigennem strømmen af stoffer fra en celle til en anden menes at forekomme.
Det er dem, der kan dannes de novo mellem to cellevægge uafhængigt af cytokinese, det vil sige uden behov for en celledelingshændelse. Sekundære plasmodesmata anses for at have specielle funktionelle og strukturelle egenskaber.
Sekundære plasmodesmata dannes ved fusion af modsatte ender af allerede eksisterende "halvdele" af plasmodesmata, som sædvanligvis etableres i regioner af cellevæggen, der er blevet udtyndet. Hver fusionshalvdel skaber de centrale hulrum i en plasmodesmus.
De centrale tråde i denne type plasmodesme tilføjes senere ved passiv "indkapsling" af endoplasmatiske reticulum tubuli, og den resulterende morfologi svarer meget til den primære plasmodesmata..
Eksperter på området antyder, at der dannes sekundære plasmodesmata i celler, der gennemgår omfattende vækstprocesser (forlængelse), dvs. mellem længdecellevægge, for at kompensere for den progressive ”fortynding” af antallet af plasmodesmata, der kan opstå takket være vækst.
Plasmodesmata repræsenterer en af de vigtigste celle-celle kommunikationsveje i plantevæv. Disse strukturer tilbyder også en kanal til elektrisk signalering, til diffusion af lipider og små opløselige molekyler og endda til udveksling af transkriptionsfaktorer og makromolekyler, såsom proteiner og nukleinsyrer..
Disse kommunikationsveje tilvejebragt af plasmodesmata synes at have en væsentlig funktion i programmeringen af planteudvikling og også i koordineringen af den fysiologiske funktion af en moden plante..
De deltager i reguleringen af frigivelsen af vigtige molekyler fra det fysiologiske og udviklingsmæssige synspunkt mod floemet (som bærer saften); de griber ind i den fysiske isolering af nogle celler og væv under udvikling, hvorfor de siges at koordinere vækst, udvikling og forsvar mod patogener.
Efter invasion af en patogen svamp er plasmodesmata også involveret, da de svarer til den vigtigste intracellulære eller simplistiske invasionsti i plantevæv..

Endnu ingen kommentarer