
Det homologe kromosomer af et individ er de kromosomer, der er en del af det samme par i en diploid organisme. I biologi henviser homologi til slægtskab, lighed og / eller funktion efter fælles oprindelse.
Hvert medlem af det homologe par har en fælles oprindelse, og de findes i den samme organisme ved fusion af kønsceller. Alle kromosomer i en organisme er somatiske kromosomer, undtagen dem fra det seksuelle par.
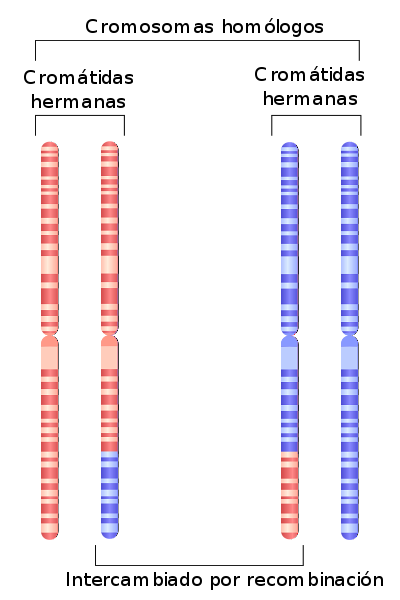
Sexkromosomer, set fra homologiens synspunkt, er en undtagelse. Begge kan have en anden oprindelse, men har regioner med homologi, der får dem til at opføre sig som somatiske kromosomer under celledelingscyklusser..
Disse homologe dele tillader både at parre sig under mitose og meiose og at rekombinere under den anden af dem..
Naturligvis er bestemte kromosompar af forskellige nært beslægtede arter også fylogenetisk homologe. Imidlertid har de rekombineret og ændret sig så meget, at det er meget vanskeligt for de samme kromosomer fra forskellige arter at være fuldstændig homologe..
Når man sammenligner kromosomerne af to arter, er homologien sandsynligvis en mosaik. Det vil sige, et kromosom af en art vil dele store eller små homologe regioner med forskellige kromosomer af den anden..
Artikelindeks
Mutationer på kromosonniveau kan opleves på to hovedniveauer: ændringer i antal og ændringer i struktur..
Ændringer på sekvensniveau analyseres på genet (og genom) og giver os en idé om ligheden i informationsindhold mellem gener, genomer og arter.
Ændringer i antal og struktur giver os mulighed for at vise ligheder og forskelle på organisatorisk niveau, enten ved at analysere individuelle kromosomer eller alle som en helhed..
Ændringer i antallet af en persons kromosomer, der påvirker en eller et par kromosomer, kaldes aneuploidier. For eksempel siges et individ med 3 kromosomer 21 i stedet for to at have en trisomi.
En trisomi på kromosom 21 er den mest almindelige årsag til Downs syndrom. På den anden side er en kvinde af den menneskelige art med et enkelt X-kromosom også aneuploid for det kromosom. XO-kvinder har det såkaldte Turners syndrom.
Ændringer, der påvirker det grundlæggende antal kromosomer i en art kaldes euploidier. Det vil sige, der er en gentagelse af det haploide kromosomsæt af arten.
Hvis der er to, er organismen diploid - som det er tilfældet med de fleste arter, der udviser seksuel reproduktion. Hvis de præsenterer tre, er organismen triploid; hvis fire, tetraploide og så videre.
Dette er meget almindeligt i planter og har været en vigtig kilde til evolutionære ændringer i denne gruppe af organismer..
Individuelle kromosomer kan også præsentere visse typer omlejringer, der kan have store konsekvenser for både individet og arten. Disse ændringer inkluderer sletninger, indsættelser, translokationer, fusioner og inversioner..
Ved sletninger går dele af kromosomet fuldstændigt tabt, hvilket giver anledning til ændringer i de meiotiske delingscyklusser med den deraf følgende produktion af muligvis ikke-levedygtige gameter..
Manglen på regioner med homologi er årsagen til unormale rekombinationshændelser. Det samme sker i tilfælde af indsættelse, da udseendet af regioner i et og ikke et andet kromosom har den samme effekt i dannelsen af regioner, der ikke er helt homologe..
En særlig tilføjelse er duplikering. I dette tilfælde føjes en del af DNA'et, der genereres i kromosomet, til en region af kromosomet. Det vil sige, det kopieres og indsættes ved siden af kopikilden.
I kromosomernes evolutionære historie har batchdubleringer spillet en grundlæggende rolle i definitionen af centromere regioner..
En anden måde at delvist ændre homologien mellem to kromosomer er ved udseendet af inverterede regioner. Oplysningerne om det omvendte område er de samme, men dets retning er modsat den for det andet parmedlem..
Dette tvinger de homologe kromosomer til at parre sig unormalt, hvilket giver anledning til andre typer yderligere omlægninger i kønscellerne. De meiotiske produkter fra disse meioser er muligvis ikke levedygtige.
En hel kromosomregion kan migrere fra et kromosom til et andet i en begivenhed kaldet en translokation. Interessant kan translokationer fremmes af stærkt konserverede regioner mellem kromosomer, der ikke nødvendigvis er homologe. Endelig er der også muligheden for at observere fusioner mellem kromosomer.
Sythenia henviser til graden af konservering af rækkefølgen af gener, når to eller flere kromosomer eller forskellige genomiske eller genetiske regioner sammenlignes.
Synthenia er ikke optaget af at studere eller måle graden af sekvenslighed mellem homologe regioner. Snarere at katalogisere informationsindholdet i disse regioner og analysere, om de er organiseret på samme måde i det rum, de optager..
Alle de omlejringer, som vi har nævnt ovenfor, mindsker naturligvis synteten mellem det ændrede kromosom og dets modstykke. De er stadig homologe, fordi de har samme oprindelse, men graden af synthenia er meget lavere.
Synthenia er nyttig til analyse af fylogenetiske forhold mellem arter. Det bruges også til at spore evolutionære baner og til at estimere den vægt, som kromosomale omlejringer har spillet i arternes udseende. Da det gør brug af store regioner, er dette makrosynteniestudier.
Microsyntenia beskæftiger sig på den anden side med at udføre den samme type analyse, men i mindre regioner, generelt på gen- eller gener-niveau. Gener såvel som kromosomer kan også gennemgå inversioner, sletninger, fusioner og tilføjelser..
Hvis de er homologe, skal to regioner af DNA have høj lighed på sekvensniveauet. Under alle omstændigheder er vi her interesserede i at påpege, at homologi er et absolut udtryk: man er homolog eller ej. Ligheden er derimod målelig.
Derfor kan to gener, der koder for den samme ting i to forskellige arter, på sekvensniveau præsentere en lighed på for eksempel 92%.
Men at sige, at begge gener er 92% homologe, er en af de værste konceptuelle fejl, der kan eksistere på det biologiske niveau..

Endnu ingen kommentarer