
Det endoplasmatisk retikulum det er en membranøs cellulær organel, der er til stede i alle eukaryote celler. Dette komplekse system optager ca. mere end halvdelen af membranerne i en fælles dyrecelle. Membranerne fortsætter, indtil de møder kernemembranen og danner et kontinuerligt element.
Denne struktur er fordelt gennem cellecytoplasmaet i form af en labyrint. Det er et slags netværk af tubuli forbundet med hinanden med sac-lignende strukturer. Inden for det endoplasmatiske retikulum forekommer protein og lipidbiosyntese. Næsten alle proteiner, der skal bæres til ydersiden af cellen, passerer først gennem reticulum.
Reticulummembranen er ikke kun ansvarlig for at adskille det indre af denne organel fra det cytoplasmatiske rum og formidle transporten af molekyler mellem disse cellekamre; Det er også involveret i syntesen af lipider, som vil udgøre en del af plasmamembranen i cellen og membranerne i de andre organeller..
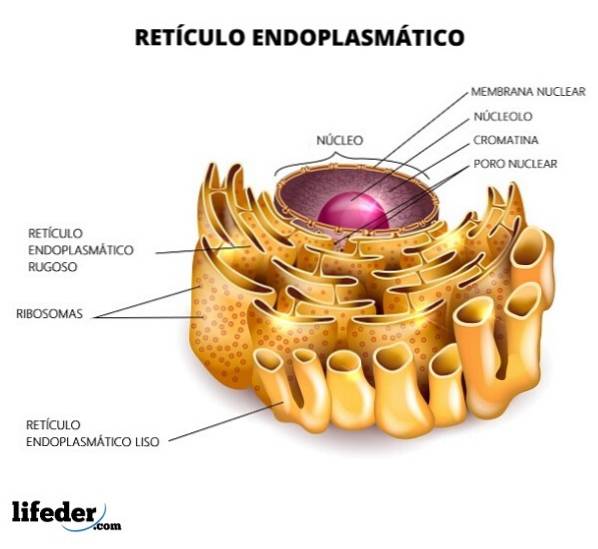
Reticulum er opdelt i glat og ru, afhængigt af tilstedeværelsen eller fraværet af ribosomer i dets membraner. Det ru endoplasmatiske retikulum har ribosomer fastgjort til membranen (tilstedeværelsen af ribosomer giver det et "groft" udseende), og tubulernes form er lidt lige.
På den anden side mangler det glatte endoplasmatiske retikulum ribosomer, og strukturens form er meget mere uregelmæssig. Funktionen af det grove endoplasmatiske retikulum er hovedsageligt rettet mod forarbejdning af proteiner. I modsætning hertil er glat ansvarlig for lipidmetabolisme.
Artikelindeks
Det endoplasmatiske retikulum er et membranøst netværk til stede i alle eukaryote celler. Den består af saccules eller cisterner og rørformede strukturer, der danner et kontinuum med kernen og er fordelt gennem cellen.
Retikulumlumen er kendetegnet ved høje koncentrationer af calciumioner ud over et oxiderende miljø. Begge egenskaber gør det muligt for den at udføre sine funktioner.
Det endoplasmatiske retikulum betragtes som den største organel, der er til stede i celler. Cellevolumenet i dette rum dækker ca. 10% af celleindretningen.
Det ru endoplasmatiske retikulum har en høj tæthed af ribosomer på overfladen. Det er regionen, hvor alle processer relateret til proteinsyntese og modifikation forekommer. Dens udseende er hovedsageligt rørformet.
Det glatte endoplasmatiske retikulum har ikke ribosomer. Det er rigeligt i celletyper, der har et aktivt stofskifte i lipidsyntese; for eksempel i cellerne i testiklerne og æggestokkene, som er steroidproducerende celler.
Tilsvarende findes det glatte endoplasmatiske retikulum i en forholdsvis høj andel i leverceller (hepatocytter). Produktionen af lipoproteiner sker i dette område..
Sammenlignet med det grove endoplasmatiske retikulum er dets struktur mere kompliceret. Overfladen af glat versus groft retikulum afhænger primært af celletypen og dens funktion..
Den fysiske arkitektur af det endoplasmatiske retikulum er et kontinuerligt system af membraner, der består af indbyrdes forbundne sække og tubuli. Disse membraner strækker sig til kernen og danner et enkelt lumen.
Gitteret er bygget af flere domæner. Fordelingen er forbundet med andre organeller, forskellige proteiner og komponenter i cytoskelettet. Disse interaktioner er dynamiske.
Strukturelt består det endoplasmatiske retikulum af den nukleare hylster og det perifere endoplasmatiske retikulum, der består af tubuli og sække. Hver struktur er relateret til en bestemt funktion.
Den nukleare hylster består, som alle biologiske membraner, af et lipid dobbeltlag. Det indre afgrænset af dette deles med det perifere retikulum.
Sækkene, der udgør det endoplasmatiske retikulum, er flade og ofte stablet. De indeholder buede områder ved membranernes kanter. Det rørformede netværk er ikke en statisk enhed; kan vokse og omstrukturere.
Sækken og tubuli-systemet er til stede i alle eukaryote celler. Det varierer dog i form og struktur afhængigt af celletypen..
Retikulum af celler med vigtige funktioner i proteinsyntese består primært af sække, mens de celler, der er mest relateret til lipidsyntese og calciumsignalering, består af et større antal tubuli..
Eksempler på celler med et stort antal sække er de sekretoriske celler i bugspytkirtlen og B-cellerne. I modsætning hertil har muskelceller og leverceller et netværk af fremtrædende tubuli..
Det endoplasmatiske retikulum er involveret i en række processer, herunder proteinsyntese, handel og foldning og modifikationer, såsom dannelse af disulfidbinding, glycosylering og tilsætning af glycolipider. Derudover deltager det i biosyntese af membranlipider.
Nylige undersøgelser har relateret retikulumet til reaktioner på cellulær stress og kan endda inducere apoptoseprocesser, selvom mekanismerne ikke er blevet fuldstændigt belyst. Alle disse processer er beskrevet i detaljer nedenfor:
Det endoplasmatiske retikulum er tæt knyttet til proteinhandel; specifikt til proteiner, der skal sendes til udlandet, til Golgi-apparatet, til lysosomer, til plasmamembranen og logisk til dem, der hører til det samme endoplasmatiske retikulum.
Det endoplasmatiske retikulum er den cellulære adfærd, der er involveret i syntesen af proteiner, der skal bæres uden for cellen. Denne funktion blev belyst af en gruppe forskere i 1960'erne og studerede celler i bugspytkirtlen, hvis funktion er at udskille fordøjelsesenzymer..
Denne gruppe, ledet af George Palade, formåede at mærke proteiner ved hjælp af radioaktive aminosyrer. På denne måde var det muligt at spore og lokalisere proteinerne ved hjælp af en teknik kaldet autoradiografi..
Radioaktivt mærkede proteiner kunne spores til det endoplasmatiske retikulum. Dette resultat indikerer, at retikulum er involveret i syntesen af proteiner, hvis endelige destination er sekretion..
Senere flytter proteinerne til Golgi-apparatet, hvor de "pakkes" ind i vesikler, hvis indhold vil blive udskilt..
Sekretionsprocessen opstår, fordi vesikulærmembranen kan smelte sammen med plasmamembranen i cellen (begge er af lipid natur). På denne måde kan indholdet frigives til ydersiden af cellen..
Med andre ord skal secernerede proteiner (og også proteiner målrettet mod lysosomer og plasmamembranen) følge en bestemt vej, der involverer det ru endoplasmatiske retikulum, Golgi-apparatet, sekretoriske vesikler og endelig det ydre af cellen..
Proteiner, der er bestemt til at blive inkorporeret i noget biomembran (plasmamembran, Golgi-membran, lysosom eller reticulum) indsættes først i reticulummembranen og frigives ikke øjeblikkeligt i lumen. De skal følge den samme vej for sekretoriske proteiner.
Disse proteiner kan placeres i membranerne ved hjælp af en hydrofob sektor. Denne region har en række på 20 til 25 hydrobofiske aminosyrer, som kan interagere med carbonkæderne i phospholipider. Den måde, hvorpå disse proteiner indsættes, er imidlertid variabel..
Mange proteiner krydser membranen kun en gang, mens andre gør det gentagne gange. Ligeledes kan det i nogle tilfælde være enden af carboxylterminalen eller aminoterminalen.
Orienteringen af proteinet etableres, mens peptidet vokser og overføres til det endoplasmatiske retikulum. Alle proteindomæner, der peger mod retikulumlumen, findes på ydersiden af cellen i deres endelige placering..
Molekyler af proteinkarakter har en tredimensionel konformation, der er nødvendig for at udføre alle deres funktioner..
DNA (deoxyribonukleinsyre), ved en proces kaldet transkription, overfører sin information til et RNA (ribonukleinsyre) molekyle. RNA passerer derefter ind i proteinerne gennem translationen. Peptider overføres til retikulum, når oversættelsesprocessen er i gang.
Disse kæder af aminosyrer er arrangeret på en tredimensionel måde inden i retikulum ved hjælp af proteiner kaldet chaperoner: et protein fra Hsp70-familien (varmechokproteiner eller varmechokproteiner for dets akronym på engelsk; tallet 70 refererer til dets atommasse, 70 KDa) kaldet BiP.
BiP-proteinet kan binde til polypeptidkæden og formidle dets foldning. Ligeledes deltager det i samlingen af de forskellige underenheder, der udgør den kvaternære struktur af proteiner..
Proteiner, der ikke er korrekt foldet, bibeholdes af retikulumet og forbliver bundet til BiP eller nedbrydes.
Når cellen udsættes for stressbetingelser, reagerer retikulumet på det, og som følge heraf forekommer den korrekte foldning af proteiner ikke. Cellen kan henvende sig til andre systemer og producere proteiner, der opretholder reticulum-homeostase.
En disulfidbro er en kovalent binding mellem sulfhydrylgrupperne, der er en del af aminosyrestrukturen cystein. Denne interaktion er afgørende for, at visse proteiner fungerer; ligeledes definerer den strukturen af proteinerne, der præsenterer dem.
Disse bindinger kan ikke dannes i andre celleområder (for eksempel i cytosolen), fordi den ikke har et oxiderende miljø, der favoriserer dets dannelse..
Der er et enzym involveret i dannelsen (og brydningen) af disse bindinger: proteindisulfidisomerasen.
I retikulumet forekommer glycosyleringsprocessen ved specifikke asparaginrester. Ligesom proteinfoldning sker glykosylering, mens translationsprocessen kører.
Oligosaccharidenhederne består af fjorten sukkerrester. De overføres til asparagin med et enzym kaldet oligosacaryltransferase, der er placeret i membranen.
Mens proteinet er i retikulum, fjernes tre glukoserester og en mannoserest. Disse proteiner føres til Golgi-apparatet til yderligere behandling..
På den anden side er visse proteiner ikke forankret til plasmamembranen af en del af hydrofobe peptider. I modsætning hertil er de bundet til visse glycolipider, der fungerer som et forankringssystem og kaldes glycosylphosphatidylinositol (forkortet GPI)..
Dette system er samlet i reticulummembranen og involverer binding af GPI ved proteinets terminale carbon..
Det endoplasmatiske retikulum spiller en afgørende rolle i lipidbiosyntese; specifikt det glatte endoplasmatiske retikulum. Lipider er en uundværlig komponent i plasmamembranerne i celler.
Lipider er stærkt hydrofobe molekyler, så de kan ikke syntetiseres i vandige omgivelser. Derfor forekommer dens syntese i forbindelse med allerede eksisterende membranagtige komponenter. Transporten af disse lipider forekommer i vesikler eller af transportørproteiner.
Membranerne i eukaryote celler består af tre typer lipider: phospholipider, glycolipider og cholesterol..
Phospholipider stammer fra glycerol og er de vigtigste strukturelle bestanddele. Disse syntetiseres i regionen af reticulummembranen, der peger på det cytosoliske ansigt. Forskellige enzymer deltager i processen.
Membranen vokser ved integration af nye lipider. Takket være eksistensen af enzymet flipase kan vækst forekomme i begge halvdele af membranen. Dette enzym er ansvarlig for at overføre lipider fra den ene side af dobbeltlaget til den anden..
Processerne med syntese af kolesterol og ceramider forekommer også i reticulum. Sidstnævnte rejser til Golgi-apparatet for at producere glycolipider eller sfingomyelin..
Calciummolekylet deltager som signalizer i forskellige processer, hvad enten det er fusion eller associering af proteiner med andre proteiner eller med nukleinsyrer..
Det indre af det endoplasmatiske retikulum har calciumkoncentrationer på 100-800 uM. Calciumkanaler og receptorer, der frigiver calcium, findes i reticulum. Calciumfrigivelse opstår, når phospholipase C stimuleres ved aktivering af G-proteinkoblede receptorer (GPCR'er).
Endvidere forekommer eliminering af phosphatidylinositol 4,5 bisphosphat i diacylglycerol og inositoltriphosphat; sidstnævnte er ansvarlig for frigivelsen af calcium.
Muskelceller har et endoplasmatisk reticulum, der er specialiseret i sekvestrering af calciumioner, kaldet det sarcoplasmatiske reticulum. Det er involveret i processerne for muskelsammentrækning og afslapning.



Endnu ingen kommentarer