
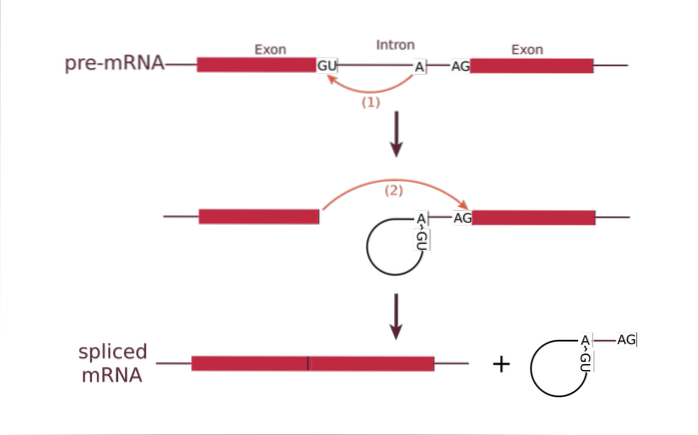
Det splejsning, eller RNA-splejsningsproces, er et fænomen, der forekommer i eukaryote organismer efter transkription af DNA til RNA og involverer eliminering af intronerne i et gen, hvorved eksoner bevares. Det betragtes som vigtigt i genekspression.
Det sker gennem begivenheder med eliminering af phosphodiesterbinding mellem exoner og introner og den efterfølgende forening af båndet mellem exoner. Splejsning forekommer i alle typer RNA, men det er mere relevant i messenger-RNA-molekylet. Det kan også forekomme i DNA- og proteinmolekyler.
Det kan være, at de på tidspunktet for samling af eksoner gennemgår et arrangement eller enhver form for ændring. Denne begivenhed er kendt som alternativ splejsning og har vigtige biologiske konsekvenser..
Artikelindeks
Et gen er en DNA-sekvens med den information, der er nødvendig for at udtrykke en fænotype. Begrebet gen er ikke strengt begrænset til DNA-sekvenser, der udtrykkes som proteiner.
Den centrale "dogme" i biologien involverer processen med at transkribere DNA til et mellemliggende molekyle, messenger RNA. Dette oversættes igen til proteiner ved hjælp af ribosomer.
Imidlertid afbrydes i eukaryote organismer disse lange gensekvenser af en type sekvens, der ikke er nødvendig for det pågældende gen: introner. For at messenger-RNA kan oversættes effektivt, skal disse introner fjernes..
RNA-splejsning er en mekanisme, der involverer forskellige kemiske reaktioner, der bruges til at fjerne elementer, der forstyrrer sekvensen af et bestemt gen. De bevarede elementer kaldes eksoner.
Spliceosomet er et enormt proteinkompleks, der er ansvarlig for at katalysere splejsningstrinnene. Den består af fem typer små nukleare RNA'er kaldet U1, U2, U4, U5 og U6, ud over en række proteiner.
Det spekuleres i, at spliceren deltager i foldningen af præ-mRNA for at justere det korrekt med de to regioner, hvor splejsningsprocessen vil forekomme.
Dette kompleks er i stand til at genkende den konsensussekvens, som de fleste introner har nær deres 5 'og 3' ender. Det skal bemærkes, at gener er fundet i Metazoans, der ikke har disse sekvenser og bruger en anden gruppe af små nukleare RNA'er til deres genkendelse..
I litteraturen anvendes udtrykket splejsning normalt på den proces, der involverer messenger-RNA. Der er dog forskellige splejsningsprocesser, der forekommer i andre vigtige biomolekyler..
Proteiner kan også gennemgå splejsning, i dette tilfælde er det en aminosyresekvens, der fjernes fra molekylet.
Det fjernede fragment kaldes "intein". Denne proces forekommer naturligt i organismer. Molekylærbiologi har formået at skabe forskellige teknikker ved hjælp af dette princip, der involverer manipulation af proteiner.
På samme måde forekommer splejsning også på DNA-niveau. Således er to DNA-molekyler, der tidligere var adskilt, i stand til at forbinde ved hjælp af kovalente bindinger.
På den anden side er der, afhængigt af typen af RNA, forskellige kemiske strategier, hvor genet kan slippe af med introner. Især splejsning af præ-mRNA er en kompliceret proces, da den involverer en række trin katalyseret af spliceosomet. Kemisk forekommer processen ved transesterificeringsreaktioner.
I gær begynder for eksempel processen med spaltning af 5'-regionen ved genkendelsesstedet, intron-exon "loop" dannes gennem en 2'-5 'phosphodiesterbinding. Processen fortsætter med dannelsen af et hul i 3'-regionen, og til sidst opstår foreningen af de to eksoner.
Nogle af intronerne, der forstyrrer de nukleare og mitokondrielle gener, kan splejses uden behov for enzymer eller energi, men gennem transesterificeringsreaktioner. Dette fænomen blev observeret i kroppen Tetrahymena thermophila.
I modsætning hertil hører de fleste nukleare gener til gruppen af introner, der har brug for maskiner til at katalysere fjernelsesprocessen..
Hos mennesker er det blevet rapporteret, at der er omkring 90.000 forskellige proteiner, og det blev tidligere antaget, at der skulle være et identisk antal gener.
Med ankomsten af nye teknologier og det menneskelige genom-projekt var det muligt at konkludere, at vi kun har omkring 25.000 gener. Så hvordan er det muligt, at vi har så meget protein?
Eksonerne samles muligvis ikke i samme rækkefølge, som de blev transkriberet til RNA, men de kan arrangeres ved at etablere nye kombinationer. Dette fænomen er kendt som alternativ splejsning. Af denne grund kan et enkelt transkriberet gen producere mere end en type protein..
Denne uoverensstemmelse mellem antallet af proteiner og antallet af gener blev belyst i 1978 af forskeren Gilbert og efterlod det traditionelle koncept "for et gen er der et protein".
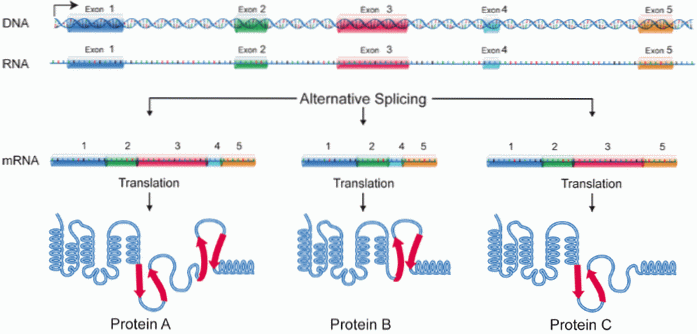
For Kelemen et al. (2013) "er en af funktionerne ved denne begivenhed at øge mangfoldigheden af messenger-RNA'er ud over at regulere forholdet mellem proteiner, mellem proteiner og nukleinsyrer og mellem proteiner og membraner."
Ifølge disse forfattere er "alternativ splejsning ansvarlig for at regulere placeringen af proteiner, deres enzymatiske egenskaber og deres interaktion med ligander". Det har også været relateret til processerne med celledifferentiering og udviklingen af organismer.
I lyset af evolutionen ser det ud til at være en vigtig mekanisme til forandring, da en høj andel af højere eukaryote organismer har vist sig at lide under høje begivenheder med alternativ splejsning. Ud over at spille en vigtig rolle i differentieringen af arter og i genomets udvikling.
Der er tegn på, at enhver fejl i disse processer kan føre til unormal cellefunktion, hvilket giver alvorlige konsekvenser for individet. Blandt disse potentielle patologier skiller kræft sig ud.
Af denne grund er alternativ splejsning blevet foreslået som en ny biologisk markør for disse unormale tilstande i celler. Ligeledes, hvis det er muligt fuldt ud at forstå grundlaget for den mekanisme, hvormed sygdommen opstår, kunne der foreslås løsninger til dem..


Endnu ingen kommentarer