Det DNA (deoxyribonukleinsyre) er det biomolekyle, der indeholder alle de oplysninger, der er nødvendige for at generere en organisme og opretholde dens funktion. Den består af enheder kaldet nukleotider, der består af en fosfatgruppe, et sukkermolekyle med fem kulstof og en nitrogenholdig base..
Der er fire nitrogenholdige baser: adenin (A), cytosin (C), guanin (G) og thymin (T). Adenin parres altid med thymin og guanin med cytosin. Beskeden indeholdt i DNA-strengen omdannes til et messenger-RNA, og dette deltager i syntesen af proteiner.

DNA er et ekstremt stabilt molekyle, negativt ladet ved fysiologisk pH, der associeres med positive proteiner (histoner) for effektivt at komprimere i kernen i eukaryote celler. En lang DNA-kæde danner sammen med forskellige tilknyttede proteiner et kromosom.
Artikelindeks
I 1953 formåede amerikaneren James Watson og den britiske Francis Crick at belyse den tredimensionelle struktur af DNA takket være det arbejde inden for krystallografi, der blev udført af Rosalind Franklin og Maurice Wilkins. De baserede også deres konklusioner på andre forfatteres arbejde.
Når DNA udsættes for røntgenstråler, dannes et diffraktionsmønster, der kan bruges til at udlede strukturen af molekylet: en helix med to antiparallelle kæder, der roterer til højre, hvor begge kæder er forbundet med hydrogenbindinger mellem baserne. . Det opnåede mønster var følgende:

Strukturen kan antages efter Braggs diffraktionslove: når en genstand er indskudt i midten af en røntgenstråle, reflekteres den, da genstandens elektroner interagerer med strålen..
Den 25. april 1953 blev resultaterne af Watson og Crick offentliggjort i det prestigefyldte magasin Natur, i en artikel på kun to sider med titlen “Molekylær struktur af nukleinsyrer”, Hvilket ville revolutionere inden for området biologi.
Takket være denne opdagelse modtog forskerne Nobelprisen i medicin i 1962 med undtagelse af Franklin, der døde før fødslen. I øjeblikket er denne opdagelse en af de store eksponenter for succesen med den videnskabelige metode til at tilegne sig ny viden.
DNA-molekylet består af nukleotider, enheder, der består af et sukker med fem carbonatomer bundet til en fosfatgruppe og en nitrogenholdig base. Den type sukker, der findes i DNA, er af deoxyribose-typen og dermed dens navn, deoxyribonukleinsyre..
For at danne kæden er nukleotiderne kovalent bundet af en phosphodiester-binding gennem en 3'-hydroxylgruppe (-OH) fra et sukker og 5'-phosphapho af det næste nukleotid.
Nukleotider bør ikke forveksles med nukleosider. Sidstnævnte refererer til den del af nukleotidet, der kun dannes af pentose (sukker) og den nitrogenholdige base.
DNA består af fire typer nitrogenholdige baser: adenin (A), cytosin (C), guanin (G) og thymin (T).
Kvælstofbaser er klassificeret i to kategorier: puriner og pyrimidiner. Den første gruppe består af en ring på fem atomer fastgjort til en anden ring på seks, mens pyrimidinerne er sammensat af en enkelt ring.
Af de nævnte baser stammer adenin og guanin fra puriner. I modsætning hertil tilhører gruppen af pyrimidiner thymin, cytosin og uracil (til stede i RNA-molekylet).
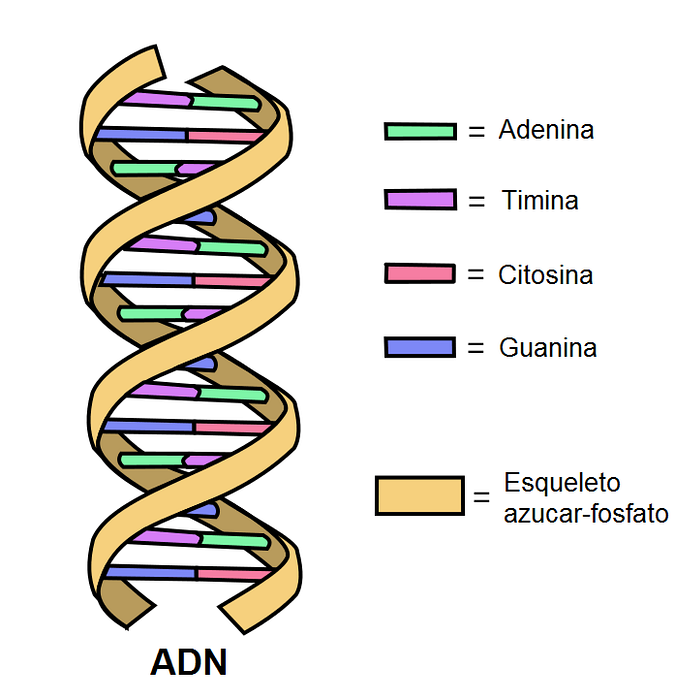
Et DNA-molekyle består af to kæder af nukleotider. Denne "kæde" er kendt som en DNA-streng..
De to tråde er forbundet med hydrogenbindinger mellem de komplementære baser. Nitrogenbaser er kovalent bundet til en rygrad af sukker og fosfater.
Hvert nukleotid placeret på en streng kan kobles med et andet specifikt nukleotid på den anden streng til dannelse af den velkendte dobbelte helix. For at danne en effektiv struktur kobles A altid med T ved hjælp af to hydrogenbindinger og G med C ved tre broer..
Hvis vi studerer proportionerne af nitrogenholdige baser i DNA, vil vi finde ud af, at mængden af A er identisk med mængden af T og den samme med G og C. Dette mønster er kendt som Chargaffs lov.
Denne parring er energisk gunstig, da den gør det muligt at bevare en lignende bredde i hele strukturen og opretholde en lignende afstand langs sukker-fosfat-rygradsmolekylet. Bemærk, at en bund af en ring passer til en af en ring.
Det foreslås, at den dobbelte helix består af 10,4 nukleotider pr. Omdrejning, adskilt af en centrum-til-centrum afstand på 3,4 nanometer. Rulningsprocessen giver anledning til dannelse af riller i strukturen, der er i stand til at observere en større og en mindre rille.
Rillerne opstår, fordi de glykosidiske bindinger i baseparret ikke er modsat hinanden med hensyn til deres diameter. Pyrimidin O-2 og purin N-3 findes i den mindre rille, mens den største rille er placeret i det modsatte område..
Hvis vi bruger analogien til en stige, består trinene af parene af komplementære baser til hinanden, mens skelettet svarer til de to gribeskinner..
Enderne af DNA-molekylet er ikke de samme, hvorfor vi taler om en “polaritet”. Den ene af dens ender, 3 ', bærer en -OH-gruppe, mens 5'-enden har den frie phosphatgruppe.
De to tråde er placeret antiparallel, hvilket betyder at de er placeret modsat deres polariteter som følger:
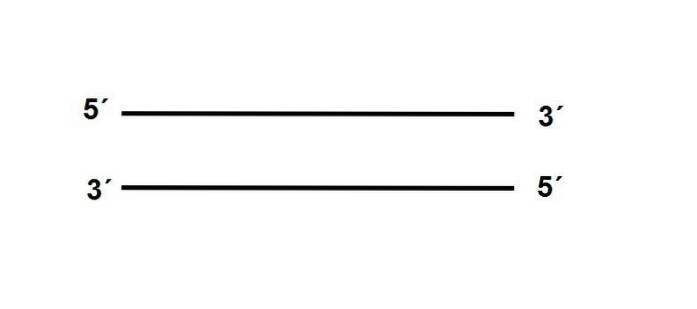
Derudover skal sekvensen af en af strengene være komplementær til dens partner, hvis den er en position er der A, i den antiparallelle streng skal der være en T.
I hver menneskelig celle er der cirka to meter DNA, der skal pakkes effektivt.
Strengen skal komprimeres, så den kan være indeholdt i en mikroskopisk kerne med en diameter på 6 um, der kun optager 10% af cellevolumenet. Dette er muligt takket være følgende komprimeringsniveauer:
I eukaryoter er der proteiner kaldet histoner, som har evnen til at binde til DNA-molekylet, hvilket er det første niveau af komprimering af strengen. Histoner har positive ladninger for at kunne interagere med de negative ladninger af DNA leveret af fosfater.
Histoner er proteiner, der er så vigtige for eukaryote organismer, at de har været praktisk talt uændrede i løbet af evolutionen - idet vi husker, at en lav grad af mutationer indikerer, at det selektive tryk på dette molekyle er stærkt. Histonskader kan føre til defekt DNA-komprimering.
Histoner kan modificeres biokemisk, og denne proces ændrer komprimeringsniveauet for det genetiske materiale.
Når histoner "hypoacetyleres" kondenseres kromatin mere, da acetylerede former neutraliserer de positive ladninger af lysiner (positivt ladede aminosyrer) i proteinet..
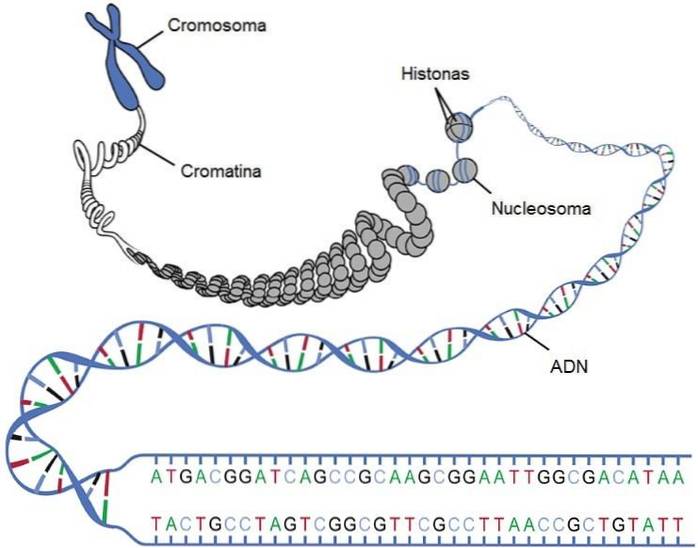
DNA-strengen vikles ind i histonerne, og de danner strukturer, der ligner perlerne på en perlekæde, kaldet nukleosomer. Kernen i denne struktur er to kopier af hver type histon: H2A, H2B, H3 og H4. Foreningen af de forskellige histoner kaldes "histonoktamer".
Oktameren er omgivet af omkring 146 basepar, der cirkler mindre end to gange. En human diploid celle indeholder ca. 6,4 x 109 nukleotider, der er organiseret i 30 millioner nukleosomer.
Organisation i nukleosomer gør det muligt at komprimere DNA til mere end en tredjedel af dets oprindelige længde.
I en proces med ekstraktion af genetisk materiale under fysiologiske forhold observeres det, at nukleosomer er arrangeret i en 30-nanometer fiber.
Kromosomer er den funktionelle arvelighedsenhed, hvis funktion er at bære et individs gener. Et gen er et segment af DNA, der indeholder informationen til at syntetisere et protein (eller en række proteiner). Der er imidlertid også gener, der koder for regulatoriske elementer, såsom RNA.
Alle humane celler (med undtagelse af kønsceller og røde blodlegemer) har to kopier af hvert kromosom, den ene arvet fra faderen og den anden fra moderen.
Kromosomer er strukturer, der består af et langt lineært stykke DNA associeret med proteinkomplekserne nævnt ovenfor. Normalt i eukaryoter er alt det genetiske materiale, der er inkluderet i kernen, opdelt i en række kromosomer.
Prokaryoter er organismer, der mangler en kerne. I disse arter er det genetiske materiale stærkt viklet sammen med alkaliske proteiner med lav molekylvægt. På denne måde komprimeres DNA'et og placeres i en central region i bakterien..
Nogle forfattere kalder ofte denne struktur "bakteriekromosom", selvom den ikke har de samme egenskaber som et eukaryot kromosom..
Ikke alle arter af organismer indeholder den samme mængde DNA. Faktisk er denne værdi meget variabel mellem arter, og der er ingen sammenhæng mellem mængden af DNA og organismenes kompleksitet. Denne modsigelse er kendt som "C-værdiparadoxet".
Den logiske begrundelse ville være at intuitere, at jo mere kompleks organismen er, jo mere DNA har den. Dette er dog ikke sandt i naturen..
For eksempel lungefiskgenomet Protopterus aethiopicus Det er 132 pg i størrelse (DNA kan kvantificeres i picogrammer = pg), mens det humane genom kun vejer 3,5 pg.
Det skal huskes, at ikke alt DNA i en organisme koder for proteiner, en stor mængde af dette er relateret til regulatoriske elementer og med de forskellige typer RNA.
Watson og Crick-modellen, udledt fra røntgendiffraktionsmønstre, er kendt som B-DNA-helixen og er den “traditionelle” og bedst kendte model. Der er dog to andre forskellige former, kaldet A-DNA og Z-DNA..
"A" -varianten drejer til højre, ligesom B-DNA, men er kortere og bredere. Denne form vises, når den relative fugtighed falder.
A-DNA roterer hvert 11. basepar, den største rille er smallere og dybere end B-DNA. Med hensyn til den mindre rille er dette mere overfladisk og bredt.
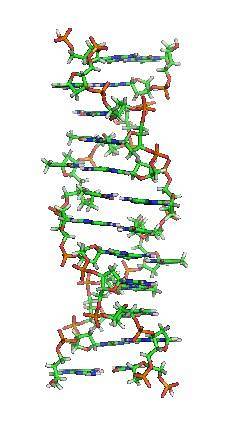
Den tredje variant er Z-DNA. Det er den smaleste form, dannet af en gruppe af hexanukleotider organiseret i en duplex af antiparallelle kæder. En af de mest fremragende egenskaber ved denne form er, at den drejer til venstre, mens de to andre måder gør det til højre..
Z-DNA vises, når der er korte sekvenser af pyrimidiner og puriner skiftevis med hinanden. Major sulcus er flad, og minor er smal og dybere sammenlignet med B-DNA.
Skønt DNA-molekylet under fysiologiske forhold hovedsagelig er i sin B-form, udsætter eksistensen af de to beskrevne varianter det genetiske materiales fleksibilitet og dynamik..
DNA-molekylet indeholder alle de oplysninger og instruktioner, der er nødvendige for konstruktionen af en organisme. Det komplette sæt genetiske oplysninger i organismer kaldes genom.
Beskeden er kodet af det "biologiske alfabet": de fire tidligere nævnte baser, A, T, G og C.
Meddelelsen kan føre til dannelse af forskellige typer proteiner eller kode for et eller andet reguleringselement. Processen, hvormed disse databaser kan levere en besked, forklares nedenfor:
Beskeden krypteret med de fire bogstaver A, T, G og C resulterer i en fænotype (ikke alle DNA-sekvenser koder for proteiner). For at opnå dette skal DNA replikere sig selv i hver celledelingsproces..
DNA-replikation er semi-konservativ: en streng tjener som skabelon til dannelsen af det nye dattermolekyle. Forskellige enzymer katalyserer replikation, herunder DNA-primase, DNA-helicase, DNA-ligase og topoisomerase..
Derefter skal meddelelsen - skrevet på et basesekvenssprog - overføres til et mellemliggende molekyle: RNA (ribonukleinsyre). Denne proces kaldes transkription..
For at transkription skal forekomme, skal forskellige enzymer deltage, herunder RNA-polymerase.
Dette enzym er ansvarlig for at kopiere DNA-beskeden og konvertere den til et messenger-RNA-molekyle. Med andre ord er transskriptionens mål at opnå messenger.
Endelig sker oversættelsen af meddelelsen til messenger-RNA-molekyler takket være ribosomerne.
Disse strukturer tager messenger-RNA'et og danner sammen med oversættelsesmaskineriet det specificerede protein..
Beskeden læses i "tripletter" eller grupper med tre bogstaver, der angiver en aminosyre - byggestenene til proteiner. Det er muligt at dechiffrere meddelelsen fra tripletterne, da den genetiske kode allerede er afsløret fuldstændigt.
Oversættelse begynder altid med aminosyren methionin, som kodes af starttripletten: AUG. "U" repræsenterer base uracil og er karakteristisk for RNA og erstatter thymin.
For eksempel, hvis messenger-RNA'et har følgende sekvens: AUG CCU CUU UUU UUA, oversættes det til følgende aminosyrer: methionin, prolin, leucin, phenylalanin og phenylalanin. Bemærk, at to tripletter - i dette tilfælde UUU og UUA - kan kode for den samme aminosyre: phenylalanin.
På grund af denne egenskab siges det, at den genetiske kode er degenereret, da en aminosyre er kodet af mere end en sekvens af tripletter, bortset fra aminosyren methionin, som dikterer starten af translationen..
Processen stoppes med specifikke stop- eller stop-tripletter: UAA, UAG og UGA. De er kendt under navnene henholdsvis okker, rav og opal. Når ribosomet registrerer dem, kan de ikke længere tilføje flere aminosyrer til kæden.
Nukleinsyrer er sure i naturen og er opløselige i vand (hydrofil). Dannelsen af hydrogenbindinger mellem phosphatgrupperne og hydroxylgrupperne af pentoser med vand kan forekomme. Det er negativt ladet ved fysiologisk pH.
DNA-opløsninger er meget tyktflydende på grund af den dobbelte spirals deformationsmodstandskapacitet, som er meget stiv. Viskositet falder, hvis nukleinsyre er enkeltstrenget.
De er meget stabile molekyler. Logisk set skal denne egenskab være uundværlig i de strukturer, der bærer genetisk information. Sammenlignet med RNA er DNA meget mere stabilt, fordi det mangler en hydroxylgruppe.
DNA kan varmedenatureres, hvilket betyder at strengene adskilles, når molekylet udsættes for høje temperaturer.
Mængden af varme, der skal tilføres, afhænger af molekylets GC-procentdel, fordi disse baser er forbundet med tre hydrogenbindinger, hvilket øger modstanden mod adskillelse..
Med hensyn til absorption af lys har de en top på 260 nanometer, hvilket øges, hvis nukleinsyren er enkeltstrenget, da nukleotidringene er eksponeret, og disse er ansvarlige for absorptionen..
Ifølge Lazcano et al. 1988 opstår DNA i overgangsfaser fra RNA og er en af de vigtigste begivenheder i livets historie.
Forfatterne foreslår tre faser: en første periode, hvor der var molekyler svarende til nukleinsyrer, senere blev genomerne sammensat af RNA, og som sidste trin dukkede dobbeltbånds-DNA-genomerne op..
Nogle beviser understøtter teorien om en primær verden baseret på RNA. For det første kan proteinsyntese forekomme i fravær af DNA, men ikke når RNA mangler. Derudover er der opdaget RNA-molekyler med katalytiske egenskaber..
Med hensyn til syntesen af deoxyribonukleotider (til stede i DNA) kommer de altid fra reduktionen af ribonukleotider (til stede i RNA).
Den evolutionære innovation af et DNA-molekyle skal have krævet tilstedeværelsen af enzymer, der syntetiserer DNA-forløbere og deltager i den omvendte transkription af RNA.
Ved at studere aktuelle enzymer kan det konkluderes, at disse proteiner har udviklet sig flere gange, og at overgangen fra RNA til DNA er mere kompleks end tidligere antaget, herunder processer til overførsel og tab af gener og ikke-ortologiske udskiftninger..
DNA-sekventering består i at belyse sekvensen af DNA-strengen med hensyn til de fire baser, der komponerer den.
Kendskab til denne sekvens er af største betydning i de biologiske videnskaber. Det kan bruges til at skelne mellem to morfologisk meget ensartede arter, til at opdage sygdomme, patologier eller parasitter og har endda en retsmedicinsk anvendelighed.
Sanger-sekventering blev udviklet i 1900'erne og er den traditionelle teknik til afklaring af en sekvens. På trods af dets alder er det en gyldig metode og i vid udstrækning brugt af forskere.
Metoden bruger DNA-polymerase, et meget pålideligt enzym, der replikerer DNA i celler, og syntetiserer en ny streng DNA ved hjælp af en allerede eksisterende som en vejledning. Enzymet kræver en først eller primer til initiering af syntese. Primeren er et lille DNA-molekyle, der er komplementært til det molekyle, der skal sekventeres.
I reaktionen tilføjes nukleotider, der vil blive inkorporeret i den nye DNA-streng af enzymet.
Ud over de "traditionelle" nukleotider inkluderer fremgangsmåden en række dideoxynukleotider for hver af baserne. De adskiller sig fra standardnukleotider i to egenskaber: strukturelt tillader de ikke, at DNA-polymerase tilføjer flere nukleotider til datterstrengen, og de har en anden fluorescerende markør for hver base.
Resultatet er en række DNA-molekyler med forskellige længder, da dideoxynukleotiderne blev inkorporeret tilfældigt og stoppet replikationsprocessen på forskellige stadier..
Denne række molekyler kan adskilles i henhold til deres længde, og nukleotidernes identitet læses ved hjælp af lysemission fra det fluorescerende mærke..
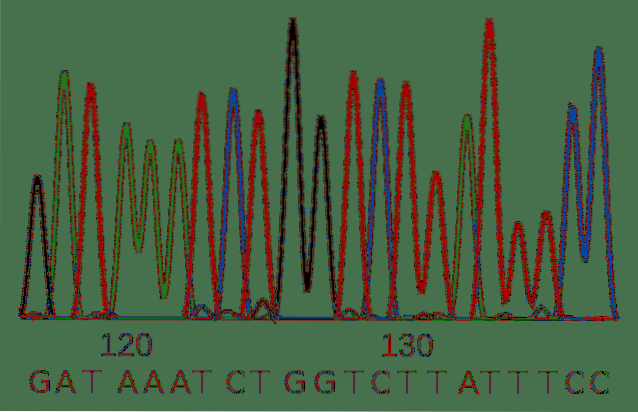
Sekventeringsteknikker udviklet i de senere år muliggør massiv analyse af millioner af prøver samtidigt.
Blandt de mest fremragende metoder er pyrosekventering, sekventering ved syntese, sekventering ved ligering og næste generations sekventering af Ion Torrent..


Endnu ingen kommentarer