Det DNA-polymerase Det er et enzym, der er ansvarlig for at katalysere polymerisationen af den nye DNA-streng under replikationen af dette molekyle. Dens hovedfunktion er at parre triphosphatdeoxyribonukleotiderne med dem i skabelonkæden. Deltager også i DNA-reparation.
Dette enzym tillader den korrekte parring mellem DNA-baserne i skabelonstrengen og den nye, efter skemaet A parrer den med T og G med C.
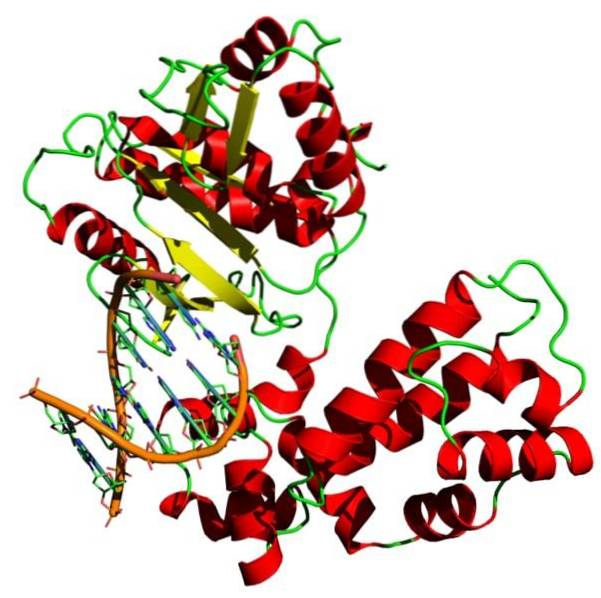
DNA-replikationsprocessen skal være effektiv og skal udføres hurtigt, så DNA-polymerase fungerer ved at tilføje ca. 700 nukleotider pr. Sekund og kun begår en fejl hver 10.9 eller 1010 inkorporerede nukleotider.
Der er forskellige typer DNA-polymerase. Disse varierer i både eukaryoter og prokaryoter, og hver har en specifik rolle i DNA-replikation og -reparation..
Det er muligt, at et af de første enzymer, der optrådte i evolution, var polymeraser, da evnen til at replikere genomet nøjagtigt er et iboende krav til udvikling af organismer..
Opdagelsen af dette enzym krediteres Arthur Kornberg og hans kolleger. Denne forsker identificerede DNA-polymerase I (Pol I) i 1956, mens han arbejdede med Escherichia coli. Tilsvarende var det Watson og Crick, der foreslog, at dette enzym kunne producere trofaste kopier af DNA-molekylet..
Artikelindeks
Prokaryote organismer (organismer uden en ægte kerne, afgrænset af en membran) besidder tre hoved-DNA-polymeraser, almindeligvis forkortet som pol I, II og III.
DNA-polymerase I deltager i DNA-replikation og reparation og har exonukleaseaktivitet i begge retninger. Dette enzyms rolle i replikation betragtes som sekundær.
II deltager i DNA-reparation, og dets exonukleaseaktivitet er i 3'-5'-forstand. III deltager i DNA-replikation og revision, og som det foregående enzym har det exonukleaseaktivitet i 3'-5'-forstand.
Eukaryoter (organismer med en ægte kerne, afgrænset af en membran) har fem DNA-polymeraser, navngivet med bogstaver i det græske alfabet: α, β, γ, δ og ε.
Polymerase γ er placeret i mitokondrierne og er ansvarlig for replikationen af genetisk materiale i denne celleorganel. I modsætning hertil findes de andre fire i cellernes kerne og er involveret i nuklear DNA-replikation..
Α-, δ- og ε-varianterne er de mest aktive i celledelingsprocessen, hvilket antyder, at deres hovedfunktion er forbundet med produktionen af DNA-kopier..
DNA-polymerase β udviser på sin side toppe af aktivitet i celler, der ikke deler sig, så det antages, at dets hovedfunktion er forbundet med DNA-reparation..
Forskellige eksperimenter har formået at verificere hypotesen om, at de for det meste forbinder α, δ og ε polymeraser med DNA-replikation. Typerne γ, δ og ε har 3'-5 'exonukleaseaktivitet.
Nye sekventeringsmetoder er lykkedes at identificere et stort udvalg af DNA-polymerasefamilier. I archaea er der specifikt identificeret en familie af enzymer, kaldet D-familien, der er unikke for denne gruppe af organismer..
DNA er molekylet, der bærer al genetisk information i en organisme. Det består af et sukker, en nitrogenholdig base (adenin, guanin, cytosin og thymin) og en phosphatgruppe.
Under processerne med celledeling, som konstant forekommer, skal DNA kopieres hurtigt og præcist - specifikt i S-fasen af cellecyklussen. Denne proces, hvor cellen kopierer DNA, er kendt som replikation.
Strukturelt består DNA-molekylet af to tråde, der danner en helix. Under replikationsprocessen adskilles disse og fungerer hver som en skabelon til dannelsen af et nyt molekyle. Således overgår de nye tråde til dattercellerne i processen med celledeling..
Da hver streng tjener som en skabelon, siges DNA-replikation at være semi-konservativ - i slutningen af processen består det nye molekyle af en ny og en gammel streng. Denne proces blev beskrevet i 1958 af forskerne Meselson og Stahl ved hjælp af isopoter.
DNA-replikation kræver en række enzymer, der katalyserer processen. Blandt disse proteinmolekyler skiller DNA-polymerase sig ud.
For at DNA-syntese skal forekomme, kræves de nødvendige substrater til processen: deoxyribonukleotidtriphosphat (dNTP)
Reaktionsmekanismen involverer et nukleofilt angreb af hydroxylgruppen i 3'-enden af den voksende streng på alfa-phosphat fra de komplementære dNTP'er, hvilket eliminerer et pyrophosphat. Dette trin er meget vigtigt, da polymerisationsenergien kommer fra hydrolysen af dNTP'erne og det resulterende pyrophosphat..
Pol III eller alfa binder til primeren (se egenskaber ved polymeraser) og begynder at tilføje nukleotider. Epsilon forlænger blykæden, og delta forlænger den forsinkede streng.
Alle kendte DNA-polymeraser har to væsentlige egenskaber forbundet med replikationsprocessen.
Først syntetiserer alle polymeraser DNA-strengen i 5'-3'-retning, idet dNTP'erne tilføjes til hydroxylgruppen i den voksende kæde..
For det andet kan DNA-polymeraser ikke begynde at syntetisere en ny streng ud af luften. De har brug for et yderligere element kendt som en primer eller primer, som er et molekyle, der består af et par nukleotider, der tilvejebringer en fri hydroxylgruppe, hvor polymerasen kan forankre sig selv og begynde sin aktivitet..
Dette er en af de grundlæggende forskelle mellem DNA- og RNA-polymeraser, da sidstnævnte er i stand til at initiere syntesen af en kæde de novo.
Den første egenskab ved DNA-polymeraser nævnt i det foregående afsnit repræsenterer en komplikation for semi-konservativ replikation. Da de to DNA-tråde kører antiparallel, syntetiseres en af dem diskontinuerligt (den, der skal syntetiseres i 3'-5'-retning).
I den forsinkede streng forekommer diskontinuerlig syntese gennem den normale aktivitet af polymerasen, 5'-3 ', og de resulterende fragmenter - kendt i litteraturen som Okazaki-fragmenter - er bundet af et andet enzym, ligase.
DNA udsættes konstant for faktorer, både endogene og eksogene, der kan skade det. Disse skader kan blokere replikering og akkumuleres, hvilket påvirker ekspressionen af gener og genererer problemer i de forskellige cellulære processer.
Ud over sin rolle i DNA-replikationsprocessen er polymerase også en nøglekomponent i DNA-reparationsmekanismer. De kan også fungere som sensorer i cellecyklussen, der forhindrer indtræden i delingsfasen, hvis DNA er beskadiget..
I øjeblikket er strukturerne af forskellige polymeraser blevet belyst takket være krystallografiske undersøgelser. Baseret på deres primære sekvens grupperes polymeraser i familier: A, B, C, X og Y.
Nogle aspekter er fælles for alle polymeraser, især dem, der er relateret til enzymets katalytiske centre.
Disse inkluderer to vigtige aktive steder, der har metalioner, med to aspartatrester og en variabel rest - enten aspartat eller glutamat, som koordinerer metallerne. Der er en anden række ladede rester, der omgiver det katalytiske centrum og konserveres i de forskellige polymeraser.
I prokaryoter er DNA-polymerase I et 103 kd polypeptid, II er et 88 kd polypeptid og III består af ti underenheder..
I eukaryoter er enzymerne større og mere komplekse: α består af fem enheder, β og γ af en underenhed, δ af to underenheder og ε af 5..
Polymerasekædereaktionen (PRC) er en metode, der anvendes i alle molekylærbiologilaboratorier takket være dens anvendelighed og enkelhed. Formålet med denne metode er at massivt amplificere et DNA-molekyle af interesse.
For at opnå dette bruger biologer en DNA-polymerase, der ikke er beskadiget af varme (høje temperaturer er afgørende for denne proces) til at amplificere molekylet. Resultatet af denne proces er et stort antal DNA-molekyler, der kan bruges til forskellige formål..
Et af de mest fremragende kliniske hjælpeprogrammer ved teknikken er dens anvendelse til medicinsk diagnose. PRC kan bruges til at kontrollere patienter for patogene bakterier og vira..
Et betydeligt antal lægemidler har til formål at afkorte mekanismerne til DNA-replikation i den patogene organisme, det være sig en virus eller en bakterie..
I noget af dette er målet inhibering af DNA-polymeraseaktivitet. For eksempel deaktiverer det kemoterapeutiske lægemiddel cytarabin, også kaldet cytosin arabinosid, DNA-polymerase.


Endnu ingen kommentarer