Det RNA eller RNA (ribonukleinsyre) er en type nukleinsyre til stede i eukaryote og prokaryote organismer og i vira. Det er en nukleotidpolymer, der indeholder fire typer nitrogenholdige baser i sin struktur: adenin, guanin, cytosin og uracil.
RNA findes generelt som et enkelt bånd (undtagen i nogle vira) på en lineær måde eller i en række komplekse strukturer. Faktisk har RNA en strukturel dynamik, der ikke observeres i DNA-dobbelthelixen. De forskellige typer RNA har meget forskellige funktioner.
Ribosomale RNA'er er en del af ribosomer, de strukturer, der er ansvarlige for proteinsyntese i celler. Messenger-RNA'er fungerer som mellemled og transporterer genetisk information til ribosomet, som oversætter meddelelsen fra en nukleotidsekvens til en aminosyresekvens..
Overførsels-RNA'er er ansvarlige for at aktivere og overføre de forskellige typer aminosyrer -20 i alt til ribosomerne. Der er et overførings-RNA-molekyle for hver aminosyre, der genkender sekvensen i messenger-RNA.
Derudover er der andre typer RNA, der ikke er direkte involveret i proteinsyntese og deltager i genregulering..
Artikelindeks
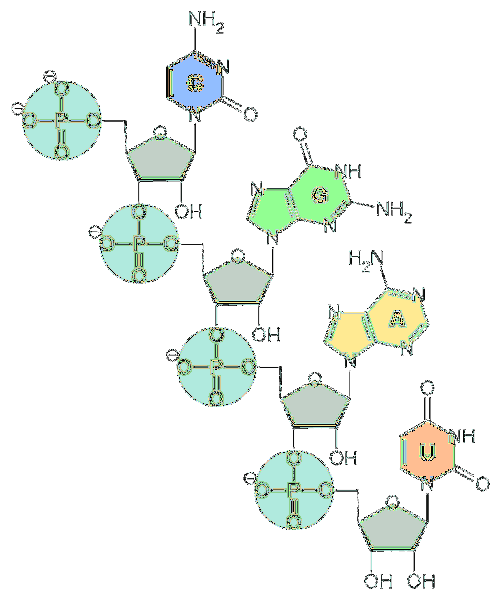
De grundlæggende enheder af RNA er nukleotider. Hvert nukleotid består af en nitrogenholdig base (adenin, guanin, cytosin og uracil), en pentose og en phosphatgruppe..
Kvælstofbaser stammer fra to grundlæggende forbindelser: pyrimidiner og puriner..
Baserne afledt af puriner er adenin og guanin, og baserne afledt af pyrimidiner er cytosin og uracil. Selvom disse er de mest almindelige baser, kan nukleinsyrer også have andre typer baser, der er mindre almindelige..
Hvad angår pentose, er de enheder af d-ribose. Af denne grund kaldes nukleotiderne, der udgør RNA, "ribonukleotider".
Nukleotider er bundet sammen ved kemiske bindinger, der involverer fosfatgruppen. For at danne dem er phosphatgruppen i 5'-enden af et nukleotid bundet til hydroxylgruppen (-OH) i 3'-enden af det næste nukleotid, hvorved der dannes en phosphodiesterlignende binding..
Langs nukleinsyrekæden har phosphodiesterbindingerne den samme orientering. Derfor er der en polaritet af strengen, der skelner mellem 3'- og 5'-enden.
Efter konvention er strukturen af nukleinsyrer repræsenteret af 5'-enden til venstre og 3'-enden til højre.
RNA-produktet fra DNA-transkription er et enkeltstrenget bånd, der drejer til højre i en spiralformet konformation ved stabling af baser. Interaktionen mellem puriner er meget større end interaktionen mellem to pyrimidiner på grund af deres størrelse.
I RNA er det ikke muligt at tale om en traditionel og reference sekundær struktur, såsom den dobbelte helix af DNA. Den tredimensionelle struktur af hvert RNA-molekyle er unik og kompleks, sammenlignelig med den for proteiner (logisk set kan vi ikke globalisere strukturen af proteiner).
Der er svage interaktioner, der bidrager til stabiliseringen af RNA, især base-stabling, hvor ringene er placeret oven på hinanden. Dette fænomen bidrager også til stabiliteten af DNA-spiralen..
Hvis RNA-molekylet finder en komplementær sekvens, vil de være i stand til at parre og danne en dobbeltstrenget struktur, der drejer til højre. Den dominerende form er type A; Med hensyn til Z-formerne er de kun påvist i laboratoriet, mens B-formen ikke er blevet observeret.
Generelt er der korte sekvenser (såsom UUGG), der er placeret i slutningen af RNA'et og har den særlige egenskab at danne sløjfer stabil. Denne sekvens deltager i foldningen af den tredimensionelle struktur af RNA.
Derudover kan der dannes hydrogenbindinger på andre steder end de typiske baseparringer (AU og CG). En af disse interaktioner forekommer mellem 2'-OH af ribose med andre grupper.
At belyse de forskellige strukturer, der findes i RNA, har tjent til at demonstrere de mange funktioner i denne nukleinsyre..
Der er to klasser af RNA: informativ og funktionel. Den første gruppe inkluderer RNA'er, der deltager i proteinsyntese og fungerer som mellemled i processen; informations-RNA'er er messenger-RNA'er.
I modsætning hertil giver RNA'er, der hører til anden klasse, de funktionelle, ikke noget nyt proteinmolekyle, og selve RNA'et er det endelige produkt. Disse er overførsels-RNA'er og ribosomale RNA'er..
I pattedyrceller er 80% af RNA ribosomalt RNA, 15% er transfer-RNA, og kun en lille del svarer til messenger-RNA. Disse tre typer arbejder sammen om at opnå proteinbiosyntese..
Der er også små nukleare RNA'er, små cytoplasmatiske RNA'er og mikroRNA'er, blandt andre. Hver af de vigtigste typer vil blive beskrevet i detaljer nedenfor:
I eukaryoter er DNA begrænset til kernen, mens proteinsyntese forekommer i cellens cytoplasma, hvor der findes ribosomer. På grund af denne rumlige adskillelse skal der være en mediator, der fører budskabet fra kernen til cytoplasmaet, og at molekylet er messenger-RNA'et..
Messenger RNA, forkortet mRNA, er et mellemliggende molekyle, der indeholder information kodet i DNA, og som specificerer en aminosyresekvens, der vil give anledning til et funktionelt protein..
Udtrykket messenger RNA blev foreslået i 1961 af François Jacob og Jacques Monod for at beskrive den del af RNA, der overførte meddelelsen fra DNA til ribosomer..
Processen med syntese af et mRNA fra DNA-strengen er kendt som transkription, og den forekommer forskelligt mellem prokaryoter og eukaryoter..
Genekspression styres af flere faktorer og afhænger af behovene i hver celle. Transskription er opdelt i tre faser: indledning, forlængelse og afslutning..
DNA-replikationsprocessen, som forekommer i hver celledeling, kopierer hele kromosomet. Transkriptionsprocessen er imidlertid meget mere selektiv, den behandler kun behandling af specifikke segmenter af DNA-strengen og kræver ikke en primer..
På Escherichia coli -den bedst studerede bakterie inden for biovidenskab - transkription begynder med afvikling af DNA-dobbelthelixen og transkriptionsløkken dannes. Enzymet RNA-polymerase er ansvarlig for syntetisering af RNA, og da transkriptionen fortsætter, vender DNA-strengen tilbage til sin oprindelige form.
Transkription initieres ikke på tilfældige steder på DNA-molekylet; der er specialiserede websteder for dette fænomen, kaldet promotorer. På E coli RNA-polymerase er koblet et par basepar over målområdet.
Sekvenserne, hvor transkriptionsfaktorer er koblet, er ret konserverede mellem forskellige arter. En af de mest kendte promotorsekvenser er TATA-boksen.
Ved forlængelse tilføjer RNA-polymeraseenzymet nye nukleotider til 3'-OH-enden efter 5 'til 3' -retningen. Hydroxylgruppen fungerer som en nukleofil og angriber alfa-phosphatet af det nukleotid, der skal tilsættes. Denne reaktion frigiver et pyrofosfat.
Kun en af DNA-strengene bruges til at syntetisere messenger-RNA'et, der kopieres i 3 'til 5'-retning (den antiparallelle form af den nye RNA-streng). Nukleotidet, der skal tilføjes, skal overholde baseparring: U-par med A og G med C.
RNA-polymerase stopper processen, når den finder regioner rige på cytosin og guanin. Til sidst er det nye messenger RNA-molekyle adskilt fra komplekset.
I prokaryoter kan et messenger-RNA-molekyle kode for mere end et protein.
Når et mRNA udelukkende koder for et protein eller polypeptid, kaldes det monocistronisk mRNA, men hvis det koder for mere end et proteinprodukt, er mRNA'et polycistronic (bemærk, at i denne sammenhæng henviser udtrykket cistron til genet).
I eukaryote organismer er langt størstedelen af mRNA'er monocistroniske, og transkriptionsmaskineriet er meget mere komplekst i denne afstamning af organismer. De er karakteriseret ved at have tre RNA-polymeraser, betegnet I, II og III, hver med specifikke funktioner.
I er ansvarlig for syntetisering af præ-rRNA'erne, II syntetiserer messenger-RNA'erne og nogle specielle RNA'er. Endelig er III ansvarlig for overførsel af RNA'er, ribosomale 5S og andre små RNA'er..
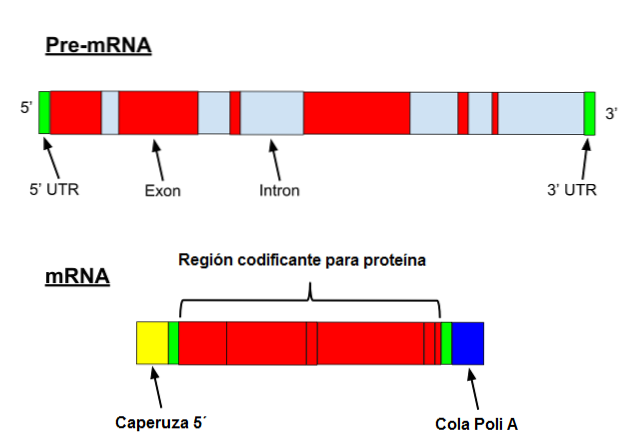
Messenger RNA gennemgår en række specifikke ændringer i eukaryoter. Den første involverer at tilføje en "cap" til 5'-enden. Kemisk er hætten en 7-methylguanosinrest bundet til enden ved hjælp af en 5 ', 5'-triphosphatbinding..
Funktionen af denne zone er at beskytte RNA mod mulig nedbrydning ved ribonukleaser (enzymer, der nedbryder RNA i mindre komponenter).
Derudover finder fjernelse af 3'-enden sted, og der tilsættes 80 til 250 adeninrester. Denne struktur er kendt som polyA "halen" og tjener som et bindingssted for forskellige proteiner. Når en prokaryote erhverver en polyA-hale, har den en tendens til at stimulere dens nedbrydning..
På den anden side transskriberes denne messenger med introner. Introner er DNA-sekvenser, der ikke er en del af genet, men som "afbryder" denne sekvens. Introner oversættes ikke og skal derfor fjernes fra messenger.
De fleste hvirveldyrgener har introner, med undtagelse af gener, der koder for histoner. Tilsvarende kan antallet af introner i et gen variere fra nogle få til snesevis af disse..
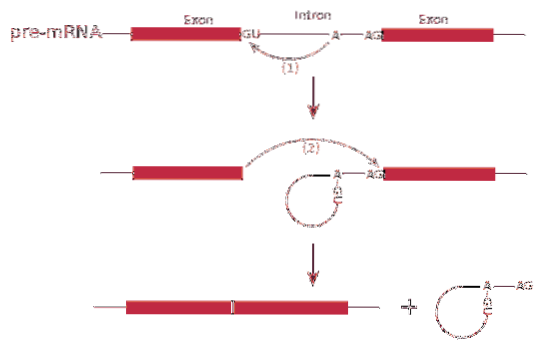
Splicing RNA eller splejsning involverer fjernelse af introner i messenger RNA.
Nogle introner, der findes i nukleare eller mitokondrie gener, kan udføre processen med splejsning uden hjælp fra enzymer eller ATP. I stedet udføres processen ved transesterificeringsreaktioner. Denne mekanisme blev opdaget i den cilierede protozo Tetrahymena thermophila.
Derimod er der en anden gruppe budbringere, der ikke er i stand til at formidle deres egne splejsning, så de har brug for yderligere maskiner. Et ret stort antal nukleare gener hører til denne gruppe..
Processen med splejsning det formidles af et proteinkompleks kaldet spliceosom eller splejsningskompleks. Systemet består af specialiserede RNA-komplekser kaldet små nukleare ribonukleoproteiner (RNP)..
Der er fem typer RNP: U1, U2, U4, U5 og U6, som findes i kernen og medierer processen med splejsning.
Det splejsning kan producere mere end en type protein - dette er kendt som splejsning alternativt, da eksoner er arrangeret differentielt, hvilket skaber sorter af messenger-RNA'er.
Ribosomalt RNA, forkortet rRNA, findes i ribosomer og deltager i proteinbiosyntese. Derfor er det en væsentlig komponent i alle celler.
Ribosomalt RNA associeres med proteinmolekyler (ca. 100) for at give anledning til ribosomale presubunits. De klassificeres afhængigt af deres sedimenteringskoefficient, betegnet med bogstavet S for Svedberg-enheder..
Et ribosom består af to dele: den store underenhed og den mindre underenhed. Begge underenheder adskiller sig mellem prokaryoter og eukaryoter med hensyn til sedimenteringskoefficienten..
Prokaryoter har en stor 50S underenhed og en lille 30S underenhed, mens i eukaryoter er den store underenhed 60S og den lille 40S.
De gener, der koder for ribosomale RNA'er, er i nucleolus, et bestemt område af kernen, der ikke er afgrænset af en membran. Ribosomale RNA'er transkriberes i denne region af RNA-polymerase I.
I celler, der syntetiserer store mængder proteiner; nucleolus er en fremtrædende struktur. Men når den pågældende celle ikke kræver et stort antal proteinprodukter, er nucleolus en næsten umærkelig struktur.
Den 60S store ribosomale underenhed er forbundet med 28S og 5.8S fragmenterne. Med hensyn til den lille underenhed (40S) er den forbundet med 18S.
I højere eukaryoter er præ-rRNA kodet i en 45S-transkriptionsenhed, som involverer RNA-polymerase I. Dette transkript behandles til modne 28S, 18S og 5.8S ribosomale RNA'er..
Efterhånden som syntese fortsætter, forbinder præ-rRNA med forskellige proteiner og danner ribonukleoproteinpartikler. Det gennemgår en række efterfølgende modifikationer, der inkluderer methylering af 2'-OH-gruppen af ribose og omdannelsen af uridinrester til pseudouridin..
Regionen, hvor disse ændringer vil forekomme, styres af mere end 150 små nukleolære RNA-molekyler, som har evnen til at parre sig til præ-rRNA..
I modsætning til resten af præ-rRNA'erne transskriberes 5S af RNA-polymerase III i nucleoplasmaet og ikke inde i nucleolus. Efter syntetisering føres det til nucleolus for at samles med 28S og 5.8S, hvorved de ribosomale enheder dannes.
Ved afslutningen af samlingsprocessen overføres underenhederne til cytoplasmaet gennem de nukleare porer..
Det kan ske, at et molekyle af messenger-RNA giver anledning til flere proteiner på samme tid, der binder til mere end et ribosom. Efterhånden som oversættelsesprocessen skrider frem, bliver messengerens afslutning fri og kan tages op af et andet ribosom, der starter en ny syntese..
Af denne grund er det almindeligt at finde ribosomer grupperet (mellem 3 og 10) i et enkelt molekyle af messenger RNA, og denne gruppe kaldes polyribosom.
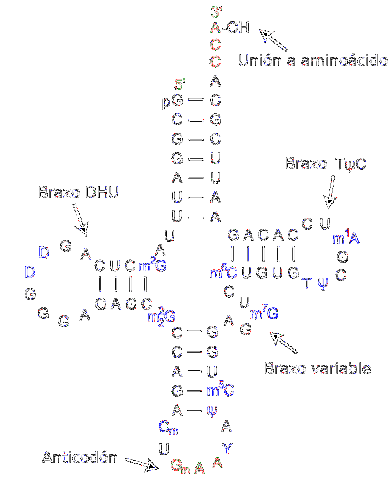
Transfer RNA er ansvarlig for overførsel af aminosyrer, når proteinsyntese processen skrider frem. De består af ca. 80 nukleotider (sammenlignet med messenger-RNA er det et "lille" molekyle).
Strukturen har folder og kryds, der minder om en trearmet klods. En adenylisk ring er placeret i en af enderne, hvor hydroxylgruppen af ribose medierer foreningen med den aminosyre, der skal transporteres.
De forskellige overførsels-RNA'er kombineres udelukkende med en af de tyve aminosyrer, der udgør proteiner; med andre ord er det køretøjet, der bærer byggestenene til proteiner. Transfer-RNA-komplekset sammen med aminosyren kaldes aminoacyl-tRNA.
Desuden genkender hver overførsels-RNA i oversættelsesprocessen - som sker takket være ribosomer - et specifikt kodon i messenger-RNA'et. Når den genkendes, frigives den tilsvarende aminosyre og bliver en del af det syntetiserede peptid.
For at genkende den type aminosyre, der skal leveres, har RNA'et et "antikodon" placeret i det midterste område af molekylet. Denne antikodon er i stand til at danne hydrogenbindinger med de komplementære baser til stede i messenger-DNA..
MicroRNA'er eller miRNA'er er en type kort, enkeltstrenget RNA mellem 21 og 23 nukleotider, hvis funktion er at regulere ekspressionen af gener. Da det ikke oversættes til protein, kaldes det ofte ikke-kodende RNA..
Ligesom andre typer RNA er mikroRNA-behandling kompleks og involverer et antal proteiner.
MicroRNA'er stammer fra længere forløbere kaldet mi-pri-RNA'er, afledt af den første transkription af genet. I cellekernen modificeres disse forløbere i mikroprocessorkomplekset, og resultatet er et præ-miRNA..
Pre-miRNA'er er 70 nukleotidhårnåle, der fortsætter deres behandling i cytoplasmaet af et enzym kaldet Dicer, som samler det RNA-inducerede dæmpningskompleks (RISC) og til sidst syntetiseres miRNA..
Disse RNA'er er i stand til at regulere ekspressionen af gener, da de er komplementære til specifikke messenger-RNA'er. Ved parring med deres mål er miRNA'erne i stand til at undertrykke budbringeren eller endog nedbryde det. Derfor kan ribosomet ikke oversætte transkriptionen..
En særlig type microRNA er små interfererende RNA'er (siRNA'er), også kaldet dæmpende RNA'er. De er korte RNA'er, mellem 20 og 25 nukleotider, som hindrer ekspressionen af visse gener.
De er meget lovende instrumenter til forskning, da de gør det muligt at lukke et gen af interesse og dermed undersøge dets mulige funktion..
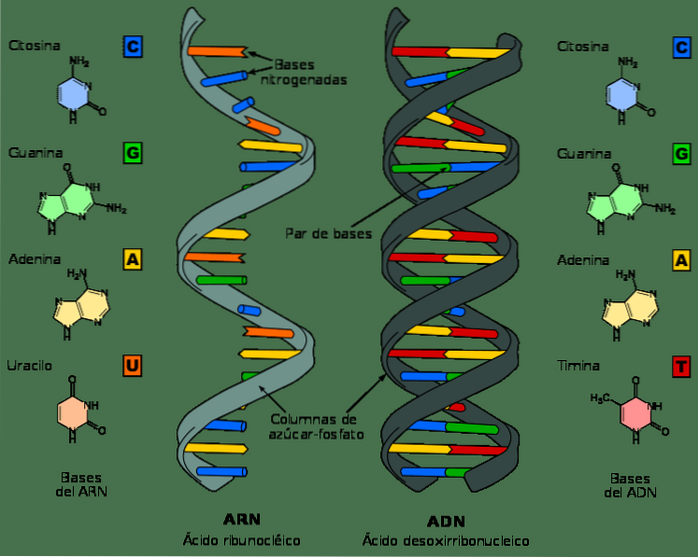
Selvom DNA og RNA er nukleinsyrer og kan se meget ens ud ved første øjekast, adskiller de sig i flere af deres kemiske og strukturelle egenskaber. DNA er et dobbeltbåndsmolekyle, mens RNA er enkeltbånd.
Som et resultat er RNA et mere alsidigt molekyle og kan tage en bred vifte af tredimensionelle former. Imidlertid har visse vira dobbeltbånds-RNA i deres genetiske materiale..
I RNA-nukleotider er sukkermolekylet en ribose, mens det i DNA er en deoxyribose, der kun adskiller sig i nærvær af et oxygenatom.
Phosphodiester-bindingen i rygraden i DNA og RNA er tilbøjelig til at gennemgå en langsom hydrolyseproces uden tilstedeværelse af enzymer. Under alkaliske forhold hydrolyserer RNA hurtigt - takket være den ekstra hydroxylgruppe - mens DNA ikke gør det.
Tilsvarende er de nitrogenholdige baser, der udgør nukleotider i DNA, guanin, adenin, thymin og cytosin; derimod erstattes thymin i RNA med uracil. Uracil kan parres med adenin på samme måde som thymin i DNA.
RNA er det eneste kendte molekyle, der er i stand til at lagre information og katalysere kemiske reaktioner på samme tid; Af denne grund foreslår flere forfattere, at RNA-molekylet var afgørende i livets oprindelse. Overraskende nok er ribosomsubstrater andre RNA-molekyler.
Opdagelsen af ribozymer førte til den biokemiske omdefinering af "enzym" - siden tidligere blev udtrykket udelukkende brugt på proteiner med katalytisk aktivitet- og hjalp med at understøtte et scenarie, hvor de første livsformer kun brugte RNA som genetisk materiale.



Endnu ingen kommentarer