Det erytroblaster de er forløberceller af hvirveldyrs erythrocytter. Faldet i iltkoncentrationen i vævene vil fremme cellulære differentieringshændelser i disse celler, der vil give anledning til modne erytrocytter. Sættet med alle disse begivenheder er kendt som erythropoiesis.
Under erythropoiesis øges hæmoglobinsyntese. Et rigeligt protein i erytrocytter, der medierer iltafgivelse til væv og afgiftning af kuldioxid fra væv, et celletoksisk affaldsprodukt fra cellulær respiration.
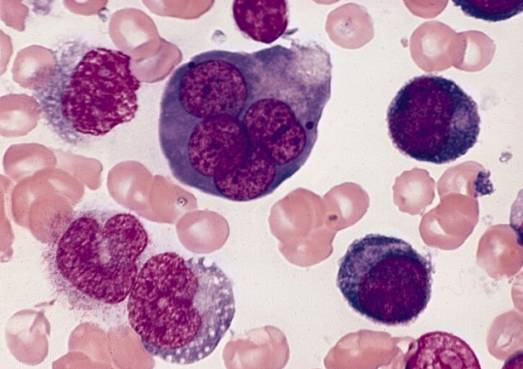
Fejl i differentieringsprocessen af erythroblaster giver anledning til et sæt blodpatologier, der sammen kaldes megaloblastiske anæmier.
Artikelindeks
De har en karakteristisk morfologi svarende til bikoncave skiver, og deres hovedfunktion er at udføre transporten af ilt (O2) til de forskellige væv i kroppen, samtidig med at det afgifter dem fra kuldioxid (CO2), der produceres under cellulær respiration..
Denne udveksling af CO2 med O2 er mulig, fordi disse celler rummer store mængder af et karakteristisk rødt protein kaldet hæmoglobin, der er i stand til at interagere med begge kemiske arter gennem en hæmgruppe, der er til stede i deres struktur..
En særlig egenskab ved disse celler i pattedyr med hensyn til resten af hvirveldyr er manglen på kerne og cytoplasmatiske organeller. I de indledende produktionsfaser i de tidlige stadier af den embryonale udvikling er det imidlertid blevet observeret, at de cellulære forløbere, hvorfra de stammer, udgør en forbigående kerne.
Sidstnævnte er ikke overraskende i betragtning af at de tidlige stadier af fostrets udvikling normalt er ens i alle hvirveldyr, hvilket kun divergerer de faser, der kompromitterer større differentiering..
Erythroblaster er celler, der vil give anledning til modne erythrocytter efter at have gennemgået på hinanden følgende begivenheder med celledifferentiering..
Disse forløberceller stammer fra en fælles myeloid stamfader i hvirveldyrs knoglemarv som kerneholdige celler forsynet med kerner og cellulære organeller..
Ændringer i indholdet af dets cytoplasma og i omlægningen af cytoskelettet vil kulminere i dannelsen af erytrocytter, der er klar til omsætning. Disse ændringer reagerer på miljømæssige stimuli, der indikerer faldet i ilt i vævene og derfor et behov i produktionen af erytrocytter..
Erythropoiesis er det udtryk, der bruges til at definere den proces, hvormed produktion og udvikling af røde blodlegemer finder sted, nødvendigt for at opretholde iltforsyningen til de forskellige organer og væv..
Denne proces reguleres fint af virkningen af erythropoietin (EPO), et nyresyntesehormon, der igen moduleres af de iltkoncentrationer, der er tilgængelige i vævene..
Oxygenkoncentrationer med lavt væv inducerer EPO-syntese ved hypoxi-inducerbar transkriptionsfaktor (HIF-1), som stimulerer erythrocytproliferation ved binding til EpoR-receptorer, der er til stede på erythrocytforløberceller.
Hos pattedyr udføres erythropoiesis i to faser, der kaldes primitiv erythropoiesis og definitiv erythropoiesis..
Den første forekommer i æggeblommesækken under den embryonale udvikling, hvilket giver anledning til store kerneholdige erytroblaster, mens den anden forekommer i føtal lever og fortsætter i knoglemarven efter anden graviditetsmåned, hvilket genererer mindre enuklerede erytrocytter..
Andre proteiner, såsom det antipoptotiske cytokin Bcl-X, hvis transkription er reguleret af transkriptionsfaktoren GATA-1, påvirker også erythropoieseprocessen positivt. Derudover er levering af jern, vitamin B12 og folinsyre også nødvendig.
Under processen med endelig erythropoiesis dannes erythrocytter i knoglemarven fra en udifferentieret stamceller eller almindelig myeloid stamfader, der er i stand til at give anledning til andre celler, såsom granulocytter, monocytter og blodplader..
Denne celle skal modtage de passende ekstracellulære signaler for at kompromittere dens differentiering mod erythroid-slægten..
Når denne forpligtelse er erhvervet, begynder en sekvens af differentieringsbegivenheder, der begynder med dannelsen af pronormoblasten, også kendt som proerythroblast. En stor erythroblast-forløbercelle med en kerne.
Derefter vil proerythroblasten opleve et progressivt fald i nuklear cellevolumen ledsaget af en stigning i hæmoglobinsyntese. Alle disse ændringer sker langsomt, når denne celle passerer gennem forskellige celletrin: den basofile erythroblast eller normoblast, polychromatisk erythroblast og ortochromatisk erythroblast.
Processen afsluttes med det samlede tab af kernen såvel som af organellerne til stede i den ortokromatiske erythroblast, der stammer fra en moden erythrocyt..
For endelig at nå dette, skal sidstnævnte passere gennem reticulocytstadiet, en enukleret celle, der stadig indeholder organeller og ribosomer i dets cytoplasma. Fuldstændig fjernelse af kernen og organellerne udføres ved exocytose.
Modne erytrocytter forlader knoglemarven i blodbanen, hvor de forbliver i cirkulation i ca. 120 dage, før de opsluges af makrofager. Derfor er erythropoiesis en proces, der sker kontinuerligt gennem en organisms liv..
Da erytoblaster udvikler sig mod fuldstændig differentiering til en moden erytrocyt, gennemgår de flere ændringer i deres cytoskelet såvel som i ekspressionen af celleadhæsionsproteiner..
Actin-mikrofilamenter depolymeriseres, og et nyt spektrinbaseret cytoskelet samles. Spectrin er et perifert membranprotein, der er placeret på det cytoplasmatiske ansigt, der interagerer med ankyrin, et protein, der formidler bindingen af cytoskelettet med det 3-transmembrane protein.
Disse ændringer i cytoskelettet og i ekspressionen af Epo-receptorer såvel som de mekanismer, der modulerer dem, er kritiske for erythroid modning..
Dette skyldes det faktum, at de formidler etableringen af interaktioner mellem erythroblaster og celler, der er til stede i knoglemarvsmikromiljøet, hvilket letter transmission af de nødvendige signaler for at begynde og afslutte differentiering..
Når differentieringen er afsluttet, opstår der nye ændringer, som favoriserer tabet af vedhæftning af cellerne til margen og deres frigivelse i blodbanen, hvor de vil udføre deres funktion..
Fejl under differentieringen af erytroblaster i knoglemarven giver anledning til forekomsten af blodpatologier, såsom megaloblastiske anæmier. Disse stammer fra mangler i tilførslen af vitamin B12 og folater, der er nødvendige for at fremme erythroblastdifferentiering..
Udtrykket megaloblastisk henviser til den store størrelse, som erythroblaster og endda erythrocytter når som et produkt af ineffektiv erythropoiesis karakteriseret ved defekt DNA-syntese.



Endnu ingen kommentarer