Det globosider De er en type sfingolipider, der hører til den heterogene familie af glycosphingolipider og er kendetegnet ved, at de i deres strukturer har en polær gruppe sammensat af komplekse strukturerede glykaner bundet til et ceramidskelet ved en B-glykosidisk binding.
De klassificeres inden for "globus" -serien af glycosphingolipider ved tilstedeværelsen af en central struktur af den generelle form Galα4Galβ4GlcβCer, og deres nomenklatur er generelt baseret på antallet og typen af sukkerrester af polarhovederne.
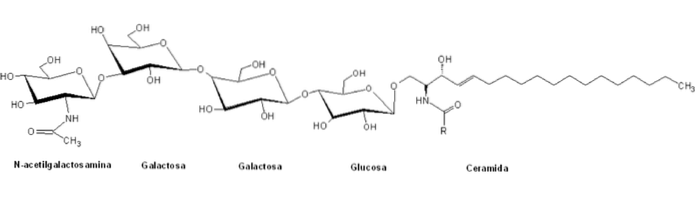
I modsætning til andre sfingolipider er globosider normale bestanddele af cellemembraner i ikke-nervøse systemiske organer hos mange pattedyr. For eksempel nyrerne, tarmene, lungerne, binyrerne og de røde blodlegemer.
Som alle membranlipider har globosider vigtige strukturelle funktioner i dannelsen og rækkefølgen af lipid-dobbeltlag..
Men i modsætning til deres sure eller phosphorylerede kolleger er globosidernes funktion ikke så meget relateret til produktionen af signalmolekyler, men snarere til deres deltagelse som en del af glykokonjugater i plasmamembranen..
Artikelindeks
De deler nogle strukturelle og funktionelle ligheder med de andre medlemmer af gruppen af glucosphingolipider: cerebrosider, gangliosider og sulfatider; herunder sammensætningen af hovedskelettet og biprodukter af dets metabolisme.
Globosider adskiller sig imidlertid fra sure glycosphingolipider (såsom gangliosider) med hensyn til ladningen af deres kulhydratpolargrupper, da de er elektrisk neutrale ved fysiologisk pH, hvilket synes at have stærke implikationer for deres funktioner som en del af den ekstracellulære matrix..
Disse polære hovedgrupper har normalt mere end to sukkermolekyler, blandt hvilke der almindeligvis er D-glucose, D-galactose og N-acetyl-D-galactosamin og i mindre grad fucose og N-acetylglucosamin.
Som med andre sfingolipider kan globosider være meget forskellige molekyler, enten under hensyntagen til de multiple kombinationer af fedtsyrer, der er bundet til sphingosin-skeletet, eller de mulige variationer af oligosaccharidkæderne i den hydrofile del..
Vejen begynder med syntesen af ceramid i det endoplasmatiske retikulum (ER). Sphingosin-skeletet dannes først ved kondensering af en L-serin og en palmitoyl-CoA.
Ceramid dannes efterfølgende ved virkningen af ceramidsyntaseenzymer, som kondenserer et andet molekyle af fedtsyre-CoA med rygraden i sfingosin ved carbonet ved position 2.
Stadig i ER kan de producerede ceramider modificeres ved tilsætning af en galactoserest til dannelse af galacto ceramider (GalCer), eller de kan i stedet transporteres til Golgi-komplekset enten ved virkningen af ceramidoverføringsproteiner (CERT) eller ved hjælp af af vesikulær transport.
I Golgi-komplekset kan ceramider glycosyleres til dannelse af glucoceramider (GlcCer).
GlcCer produceres på den cytosoliske overflade af den tidlige Golgi. Det kan derefter transporteres til kompleksets luminale overflade og derefter glykosyleres af specifikke glycosidasenzymer, der genererer mere komplekse glycosfingolipider..
De fælles forløbere for alle glycosphingolipider syntetiseres i Golgi-komplekset ved virkningen af glycosyltransferaser fra GalCer eller GlcCer.
Disse enzymer overfører specifikke kulhydrater fra de passende nukleotidsukker: UDP-glucose, UDP-galactose, CMP-sialinsyre osv..
Når GlcCer passerer gennem Golgi vesikulært menneskehandel, galaktosyleres det til at producere lactosylceramid (LacCer). LacCer er det forgreningspunkt, hvorfra forstadierne til de andre glycosphingolipider syntetiseres, dvs. molekylet, hvortil der efterfølgende tilsættes mere neutrale polære sukkerrester. Disse reaktioner katalyseres af specifikke globosidsyntaser.
Disse lipider findes hovedsageligt i humane væv. Som mange glycosphingolipider er globosider beriget på ydersiden af plasmamembranen i mange celler..
De er især vigtige i humane erytrocytter, hvor de repræsenterer den største type glycolipid på celleoverfladen..
Desuden er de, som nævnt ovenfor, en del af sættet med glykokonjugater i plasmamembranerne i mange ikke-nervøse organer, hovedsageligt nyrerne..
Globosides funktioner er ikke blevet belyst fuldt ud til dato, men det er kendt, at nogle arter øger celleproliferation og motilitet i modsætning til inhiberingen af disse begivenheder forårsaget af nogle gangliosider..
Et tetraglykosyleret globosid, Gb4 (GalNAcβ3Galα4Galβ4GlcβCer), fungerer i den stedfølsomme genkendelse af strukturelle forstyrrelser af erytrocytter under celleadhæsionsprocesser..
Nylige undersøgelser har bestemt inddragelsen af Gb4 i aktiveringen af ERK-proteiner i carcinomcellelinier, hvilket kan betyde dets deltagelse i tumorinitiering. Disse proteiner hører til den mitogenaktiverede proteinkinase (MAPK) signaleringskaskade, der består af elementerne Raf, MEK og ERK..
Deres deltagelse som receptorer for nogle bakterietoksiner fra Shiga-familien er blevet rapporteret, specifikt globosidet Gb3 (Galα4Galβ4GlcβCer), også kendt som CD77, udtrykt i umodne B-celler; også som receptorer for HIV-adhæsionsfaktor (gp120) og synes at have implikationer i visse typer kræft og andre sygdomme.
Der er mange typer lipidose hos mennesker. Globosider og deres metaboliske veje er især relateret til to sygdomme: Fabry sygdom og Sandhoff sygdom..
Det refererer til en kønsbundt nedarvet systemisk lidelse, der først ses hos patienter med flere lilla pletter i navlestrengen. Det påvirker organer som nyrer, hjerte, øjne, ekstremiteter, en del af mave-tarm-og nervesystemet.
Det er produktet af en metabolisk defekt i enzymet ceramid trihexosidase, der er ansvarlig for hydrolysen af trihexosiceramid, et mellemprodukt i katabolismen af globosider og gangliosider, der forårsager en ophobning af disse glycolipider i vævene..
Denne patologi blev oprindeligt beskrevet som en variant af Tay-Sachs sygdom relateret til metabolismen af gangliosider, men dette præsenterer også akkumuleringen af globosider i indvoldene. Det er en arvelig lidelse med autosomale recessive mønstre, der gradvis ødelægger neuroner og rygmarv.
Det har at gøre med fraværet af form A og B af enzymet β-N-acetylhexosaminidase på grund af mutationer i genet HEKS. Disse enzymer er ansvarlige for et af nedbrydningstrinnene for nogle glycosphingolipider.


Endnu ingen kommentarer