Det prometaphase det er et trin i celledelingsprocessen, mellemliggende mellem profase og metafase. Det er kendetegnet ved interaktionen mellem de delende kromosomer og de mikrotubuli, der er ansvarlige for at adskille dem. Prometaphase forekommer i både mitose og meiose, men med forskellige egenskaber.
Det klare mål med al celledeling er at producere flere celler. For at opnå dette skal cellen oprindeligt duplikere sit DNA-indhold; repliker det. Derudover skal cellen adskille disse kromosomer på en sådan måde, at det særlige formål med hver opdeling af cytoplasma er opfyldt..

I mitose er det samme antal kromosomer fra modercellen i dattercellerne. I meiose I er adskillelsen mellem homologe kromosomer. I meiose II er adskillelsen mellem søsterkromatider. I slutningen af processen opnås de fire forventede meiotiske produkter.
Cellen styrer denne komplicerede mekanisme ved brug af specialiserede komponenter såsom mikrotubuli. Disse er organiseret af centrosomet i de fleste eukaryoter. I andre, tværtimod, såsom højere planter, fungerer en anden type organisationscenter for mikrotubuli.
Artikelindeks

Mikrotubuli er lineære polymerer af proteinet tubulin. De er involveret i næsten alle cellulære processer, der involverer forskydning af en eller anden intern struktur. De er en integreret del af cytoskelet, cilia og flagella.
I tilfælde af planteceller spiller de også en rolle i den interne strukturelle organisation. I disse celler danner mikrotubuli en slags gobelin, der er fastgjort til plasmamembranens indre overflade..
Denne struktur, der kontrollerer opdeling af planteceller, er kendt som den kortikale organisering af mikrotubuli. På tidspunktet for mitotisk opdeling kollapser de for eksempel i en central ring, der vil være det fremtidige sted for den centrale plade, i det plan, hvor cellen deler sig..
Mikrotubuli er sammensat af alfa-tubulin og beta-tubulin. Disse to underenheder danner en heterodimer, som er den grundlæggende strukturelle enhed af tubulinfilamenter. Polymerisationen af dimerer fører til dannelsen af 13 protofilamenter i en lateral organisation, der giver anledning til en hul cylinder.

De hule cylindre med denne struktur er mikrotubuli, som ved deres egen sammensætning viser polaritet. Det vil sige, den ene ende kan vokse ved tilsætning af heterodimerer, mens den anden kan gennemgå subtraktioner. I sidstnævnte tilfælde krymper mikrotubuli i stedet for at forlænge i den retning.
Mikrotubuli-kerner (dvs. begynder at polymerisere) og organiseres i mikrotubuli-organiseringscentre (COM). COM er forbundet med centrosomer under opdelinger i dyreceller.
I højere planter, der ikke har centrosomer, er COM til stede på analoge steder, men består af andre komponenter. I cilia og flagella er COM placeret basalt i forhold til motorstrukturen.
Forskydningen af kromosomer under celledeling opnås gennem mikrorør. Disse formidler den fysiske interaktion mellem kromosomens centromerer og COM.
Ved målrettede depolymeriseringsreaktioner vil metafasekromosomer i sidste ende bevæge sig mod polerne i delende celler..
Korrekt mitotisk kromosomadskillelse er en, der garanterer, at hver dattercelle modtager et komplement af kromosomer, der er identiske med modercellens..
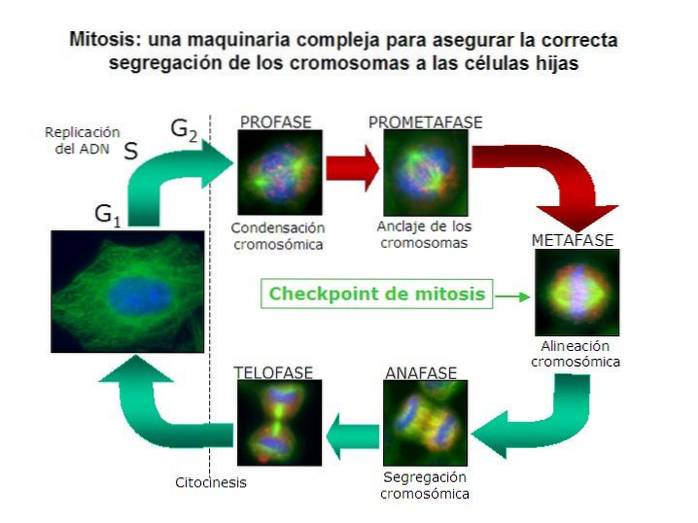
Dette betyder, at cellen skal adskille hvert par duplikerede kromosomer i to separate, individuelle kromosomer. Det vil sige, det skal adskille søsterchromatiderne for hvert homologt par fra hele komplementet af stamcellens kromosomer..
I åbne mitoser er processen med forsvinden af kernekapslen kendetegnende for prometaphase. Dette gør det muligt for den eneste hindring mellem MOC og centromererne i kromosomerne at forsvinde..
Fra MOC'erne polymeriseres lange mikrotubuli-filamenter og forlænges mod kromosomerne. Efter at have fundet et centromer ophører polymerisation, og der opnås et kromosom, der er knyttet til en COM..
I mitose er kromosomerne dobbelt. Derfor er der også to centromerer, men stadig forenet i den samme struktur. Dette betyder, at vi i slutningen af mikrotubuli-polymerisationsprocessen har to af dem pr. Duplikeret kromosom..
Et glødetråd vil vedhæfte en centromer til en COM, og en anden til søsterkromatiden, der er fastgjort til COM modsat den første..
I lukkede mitoser er processen næsten identisk med den foregående, men med en stor forskel; den nukleare konvolut forsvinder ikke. Derfor er COM intern og er forbundet med den interne nukleare konvolut gennem den nukleare lamina..
I semi-lukkede (eller semi-åbne) mitoser forsvinder kernekapslen kun på de to modsatte punkter, hvor en mitotisk COM findes uden for kernen..
Dette betyder, at mikrotubuli i disse mitoser trænger ind i kernen for at være i stand til at mobilisere kromosomerne i trin efter prometaphase..
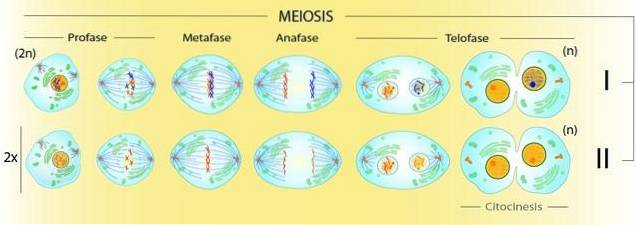
Da meiose involverer produktion af fire 'n' celler fra en '2n' celle, skal der være to divisioner af cytoplasma. Lad os se på det sådan: i slutningen af metafase I vil der være fire gange flere kromatider end centromerer synlige under mikroskopet.
Efter den første opdeling vil der være to celler med dobbelt så mange kromatider som centromerer. Først i slutningen af den anden cytoplasmatiske division vil alle centromerer og kromatider blive individualiserede. Der vil være så mange centromerer, som der er kromosomer.
Nøgleproteinet til, at disse komplekse interchromatin-interaktioner finder sted i mitose og meiose, er cohesin. Men der er flere komplikationer i meiose end i mitose. Det er derfor ikke overraskende, at meiotisk cohesin adskiller sig fra mitotisk.
Cohesins tillader sammenhængen mellem kromosomer under deres mitotiske og meiotiske kondensationsproces. Derudover tillader og regulerer de interaktionen mellem søsterkromatider i begge processer..
Men i meiose fremmer de også noget, der ikke forekommer i mitose: homolog parring og de deraf følgende synapser. Disse proteiner er forskellige i hvert tilfælde. Vi kunne sige, at meiose uden en sammenhæng, der adskiller den, ikke ville være mulig.
Mekanisk set er centromere / COM-interaktionen den samme i al celledeling. Imidlertid adskiller cellen ikke i prometaphase I af meiose I søsterkromatider, som den gør i mitose..
I modsætning hertil har den meiotiske tetrad fire kromatider i et tilsyneladende dobbelt sæt centromerer. I denne struktur er der noget andet, der ikke er til stede i mitose: chiasmata.
Kiasmer, som er fysiske forbindelser mellem homologe kromosomer, er hvad der skelner mellem hvilke centromerer der skal adskilles: de af homologe kromosomer.
Således dannes forbindelser i prometaphase I mellem homologernes centromer og COM ved modsatte poler i cellen..
Denne prometaphase II ligner mere mitotisk prometaphase end meiotisk prometaphase I. I dette tilfælde vil COM'erne "lancere" mikrotubuli ved de duplikerede centromerer af søsterkromatider..
Således vil to celler blive produceret med individuelle kromosomer, produktet af en kromatid fra hvert par. Derfor vil celler med det haploide kromosomale komplement til arten blive givet.



Endnu ingen kommentarer