Det G-proteiner eller guaninnukleotidbindende proteiner, er proteiner associeret med plasmamembranen, der hører til en familie af signal "kobler" proteiner, der har vigtige funktioner i mange signaltransduktionsprocesser i eukaryote organismer.
I litteraturen beskrives G-proteiner som binære molekylære omskiftere, da deres biologiske aktivitet bestemmes af ændringer i deres struktur givet af de nukleotidarter, som de er i stand til at binde til: guanosinnukleotider (diphosphat (BNP) og trifosfat (GTP)) ).
De aktiveres generelt af receptorer fra en familie af proteiner kendt som G-proteinkoblede receptorer (GPCR'er).G-proteinkoblede receptorer "), der modtager et eksternt initialt signal og konverterer det til konformationsændringer, der udløser aktivering, som efterfølgende oversættes til aktivering af et andet effektorprotein.
Nogle forfattere mener, at de kodende gener for denne familie af proteiner udviklede sig ved duplikering og divergens af et fælles forfædren, hvis produkt i stigende grad blev raffineret og specialiseret..
Blandt den store række af cellulære funktioner, som disse proteiner har, er translokation af makromolekyler under proteinsyntese, transduktion af hormonelle signaler og sensoriske stimuli samt regulering af celleproliferation og differentiering..
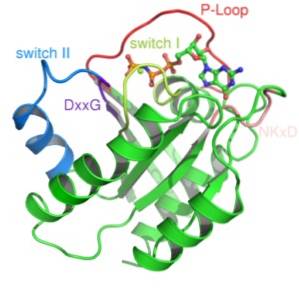
To klasser af denne type protein er blevet beskrevet: små G-proteiner og heterotrimere G-proteiner. Den første tredimensionelle struktur af et G-protein blev opnået for mere end et årti siden fra et lille G-protein kendt som Ras.
Artikelindeks
Strukturelt set genkendes to typer G-proteiner: de små G-proteiner og de meget mere komplekse heterotrimere G-proteiner..
Små G-proteiner består af et enkelt polypeptid med ca. 200 aminosyrerester og ca. 20-40 kDa, og i deres struktur er der et konserveret katalytisk domæne (G-domænet) sammensat af fem a-helixer, seks β-foldede ark og fem polypeptidsløjfer.
Heterotrimer G-proteiner er derimod integrale membranproteiner, der er sammensat af tre polypeptidkæder, kendt som α-, β- og γ-underenhederne..
-Α-underenheden vejer mellem 40 og 52 kDa, har en guaninnukleotidbindende region og har GTPase-aktivitet til hydrolysering af bindinger mellem phosphatgrupperne i GTP..
Α-underenhederne af forskellige G-proteiner deler nogle strukturelle domæner, såsom dem til binding og hydrolyse af GTP, men er meget forskellige i deres bindingssteder for receptor- og effektorproteinerne..
-Β-underenheden har en lidt lavere molekylvægt (mellem 35 og 36 kDa).
-Y-underenheden er derimod meget mindre og har en omtrentlig molekylvægt på 8 kDa..
Alle heterotrimere G-proteiner har 7 transmembrane domæner og deler sekvenslighed med β- og y-domænerne. Disse to domæner er så stærkt forbundet, at de betragtes som en enkelt funktionel enhed..
Som nævnt ovenfor er der to typer G-proteiner: små og heterotrimere..
Små G-proteiner har roller i cellevækst, proteinsekretion og intracellulær vesikelinteraktion. For deres del er heterotrimere G-proteiner associeret med transduktion af signaler fra overfladereceptorer og fungerer også som omskiftere, der veksler mellem to tilstande afhængigt af det associerede nukleotid..
Disse proteiner kaldes også små GTPaser, små GTP-bindende proteiner eller Ras-protein-superfamilie og danner en uafhængig superfamilie inden for den store klasse af GTP-hydrolaser med regulerende funktioner..
Disse proteiner er meget forskellige og styrer flere cellulære processer. De er kendetegnet ved et bevaret GTP-bindende domæne, "G" -domænet. Bindingen af dette phosphatnukleotid forårsager vigtige konformationsændringer i dets katalytiske domæne i små G-proteiner..
Dens aktivitet er tæt relateret til GTPase-aktiverende proteiner (GAP).GTPase-aktiverende protein ") og med guaninnukleotidudvekslingsfaktorer (GEF)Guaninnukleotid-udvekslingsfaktor ”).
Fem klasser eller familier af små G-proteiner er blevet beskrevet i eukaryoter:
-Ras
-Rho
-Rab
-Sar1 / Arf
-Løb
Ras- og Rho-proteinerne styrer genekspression, og Rho-proteinerne modulerer også reorganiseringen af cytoskelettet. Rab- og Sar1 / Arf-gruppeproteinerne påvirker vesikulær transport, og Ran-proteinerne regulerer nuklear transport og cellecyklussen..
Denne type protein fortjener også en tilknytning til to andre proteinfaktorer, så signalvejen fra det ydre miljø til det indre af cellen består af tre elementer i følgende rækkefølge:
Der er en stor mangfoldighed af heterotrimere G-proteiner, og dette er relateret til den store mangfoldighed af α-underenheder, der findes i naturen, hvor kun 20% af aminosyresekvensen er konserveret.
Normalt identificeres heterotrimeriske G-proteiner takket være mangfoldigheden af α-underenheden, hovedsageligt baseret på deres funktionelle og sekvensligheder..
Α-underenhederne består af fire familier (Gs-familien, Gi / o-familien, Gq-familien og G12-familien). Hver familie består af en anden “isotype”, der tilsammen tilføjer mere end 15 forskellige former for α-underenheder..
Denne familie indeholder repræsentanter, der også deltager i opreguleringen af adenylatcyclase-proteiner og udtrykkes i de fleste celletyper. Den består af to medlemmer: Gs og Golf.
Abonnementet "s " refererer til stimulering (fra engelsk “stimulering ") og abonnementet “olf " henviser til "lugt" (fra engelsk "Olfaction"). Golfproteiner udtrykkes især i sensoriske neuroner, der er ansvarlige for lugt.
Dette er den største og mest forskelligartede familie. De udtrykkes i mange celletyper og medierer receptorafhængig inhibering af forskellige typer adenylcyklusser (abonnementet "jeg " henviser til hæmning).
Proteiner med α-underenhederne i Go-gruppen udtrykkes især i celler i centralnervesystemet og har to varianter: A og B..
Proteiner med denne α-underenhedsfamilie er ansvarlige for reguleringen af phospholipase C. Denne familie består af fire medlemmer, hvis α-underenheder udtrykkes af forskellige gener. De er rigelige i leverceller, nyreceller og lunger.
Denne familie udtrykkes allestedsnærværende i organismer, og det vides ikke med sikkerhed, hvad der præcist er de cellulære processer, der reguleres gennem proteiner med disse underenheder..
Β og γ underenheder
Selv om mangfoldigheden af alfastrukturer er afgørende for identifikationen af heterotrimere proteiner, er der også en masse mangfoldighed med hensyn til de to andre underenheder: beta og gamma..
G-proteiner deltager i "kanalisering" af signaler fra receptorer på plasmamembranen til kanaler eller effektorenzymer..
Det mest almindelige eksempel på funktionen af denne type proteiner er i reguleringen af enzymet adenylatcyclase, et enzym, der er ansvarlig for syntesen af adenosin 3 ', 5'-monophosphat eller simpelthen cyklisk AMP, et molekyle, der har vigtige funktioner som anden messenger i mange kendte cellulære processer:
-Selektiv fosforylering af proteiner med specifikke funktioner
-Genetisk transkription
-Cytoskeletal reorganisering
-Sekretion
-Membranafpolarisering
De deltager også indirekte i reguleringen af inositolsignalkaskaden (phosphatidylinositol og dens phosphorylerede derivater), som er ansvarlige for styringen af calciumafhængige processer såsom kemotaxis og sekretionen af opløselige faktorer..
Mange ionkanaler og transportproteiner styres direkte af proteiner fra G-proteinfamilien, og på samme måde er disse proteiner involveret i mange sensoriske processer, såsom vision, lugt, blandt andre..
Interaktionsmåden for et G-protein med effektorproteinerne er specifik for hver klasse eller familie af proteiner.
For G-proteiner kombineret med membranreceptorer (heterotrimere G-proteiner) forårsager binding af et guaninnukleotid såsom BNP eller guanosindiphosphat til α-underenheden sammenhængen mellem de tre underenheder og danner et kompleks kendt som Gαβγ eller G-BNP, som er fastgjort til membranen.
Hvis BNP-molekylet efterfølgende udveksles med et GTP-molekyle, adskilles den α-underenhed, der er knyttet til GTP, fra β- og γ-underenhederne og danner et separat kompleks kendt som Gα-GTP, som er i stand til at ændre aktiviteten af dets enzymer eller målbærerproteiner.
Den hydrolytiske aktivitet af denne underenhed gør det muligt for den at afslutte aktiveringen, udveksle GTP med et nyt BNP og gå videre til den inaktive konformation..
I fravær af de ophidsede receptorer, der associeres med G-proteiner, er denne proces med udveksling af BNP til GTP meget langsom, hvilket betyder, at heterotrimere G-proteiner kun udveksler BNP for GTP i en fysiologisk signifikant hastighed, når de er bundet til deres exciterede receptorer.

Endnu ingen kommentarer