
To gener er forbundet når de har tendens til at blive arvet sammen som om de var en enkelt enhed. Dette kan også ske med mere end to gener. Under alle omstændigheder er denne opførsel af gener det, der har tilladt genetisk kortlægning ved binding og rekombination..
På Mendels tid havde andre forskere, såsom Boveri-ægtefællerne, observeret, at der var kroppe i cellekernen, der blev udskilt under celledeling. Disse var kromosomerne.

Senere, med Morgan og hans gruppes arbejde, var der en klarere forståelse af arv af gener og kromosomer. Det vil sige, gener adskiller sig som de kromosomer, der bærer dem (kromosomal teori om arv).
Artikelindeks
Der er, som vi ved, langt færre kromosomer end gener. Mennesket har for eksempel omkring 20.000 gener fordelt på omkring 23 forskellige kromosomer (arter af haploide belastninger).
Hvert kromosom er repræsenteret af et langt DNA-molekyle, hvor mange, mange gener er kodet separat. Hvert gen befinder sig derefter på et bestemt sted (locus) på et specifikt kromosom; til gengæld bærer hvert kromosom mange gener.
Med andre ord er alle generne på et kromosom bundet til hinanden. Hvis det ser ud til at de ikke er det, er det fordi der er en proces med fysisk udveksling af DNA mellem kromosomerne, der skaber illusionen om uafhængig distribution..
Denne proces kaldes rekombination. Hvis to gener er forbundet, men bredt adskilt fra hinanden, vil rekombination altid forekomme, og generne vil adskille sig ligesom Mendel observerede..
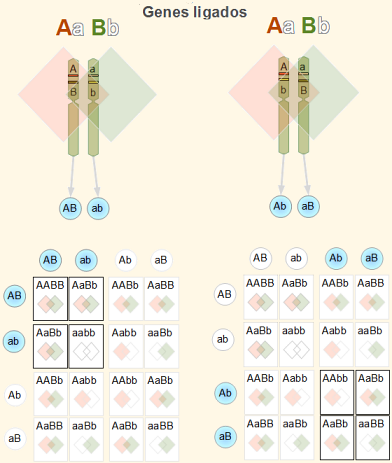
For at observere og demonstrere sammenhæng fortsætter forskeren med at krydse med individer, der præsenterer en kontrasterende manifestation af fænotypen af de gener, der undersøges (for eksempel P: AAbb x aaBB).
Alle F1-efterkommere vil være AaBb. Af dihybridkorset AaBb x aabb (eller testkors) man kunne forvente et F2 afkom, der viser de genotypiske (og fænotypiske) proportioner 1AaBb:1Aabb: 1aaBb: 1aabb.
Men dette gælder kun, hvis generne ikke er forbundet. Den første genetiske indikation af, at to gener er forbundet, er at der overvejes en overvægt af faderlige fænotyper: det vil sige, Aabb + aaBb >> AaB_b + aabb.
I tilfælde af sammenkædede gener, som vi bruger som et eksempel, producerer enkeltpersoner hovedsagelig kønsceller Ab Y aB, mere end kønsceller AB Y ab.
Da den dominerende allel i et gen er forbundet med den recessive allel i det andet gen, siges det, at begge gener er forbundet i frastødning. Hvis der observeres en overvægt af alleler AB Y ab om kønsceller Ab Y aB, gener siges at være forbundet i kobling.
Det vil sige, de dominerende alleler er koblet til det samme DNA-molekyle; eller hvad der er det samme, de er forbundet med det samme kromosom. Disse oplysninger er yderst nyttige til genetisk forbedring.
Dette gør det muligt at fastslå antallet af individer, der skal analyseres, når generne er bundet, og det ønskes at vælge for eksempel de to dominerende tegn.
Dette ville være vanskeligere at opnå, når begge gener er i frastødning, og bindingen er så tæt, at der næsten ikke er nogen rekombination mellem de to gener..
Eksistensen af selve koblingen var et stort fremskridt i vores forståelse af gener og deres organisation. Men derudover tillod det os også at forstå, hvordan selektion kan handle i befolkninger og forklare lidt udviklingen af levende væsener.
Der er gener, der er så tæt forbundne, at der kun produceres to typer gameter i stedet for de fire, der muliggør uafhængig distribution.
I ekstreme tilfælde forekommer disse to sammenkædte gener (i kobling eller frastødning) kun i en type tilknytning i befolkningen. Hvis dette sker, siges der at eksistere en ligevægtsforbindelse.
Koblingsuligevægt opstår for eksempel når manglen på de to dominerende alleler mindsker chancerne for overlevelse og reproduktion af individer..
Dette sker, når individer er et produkt af befrugtning mellem kønsceller ab. Befrugtning mellem kønsceller aB Y Ab, tværtimod øger det sandsynligheden for individets overlevelse.
Disse vil have mindst en allel TIL og en allel B, og viser de tilsvarende vilde tilknyttede funktioner.
Kobling og dens uligevægt kan også forklare, hvorfor nogle uønskede alleler af et gen ikke elimineres fra befolkningen. Hvis de er tæt knyttet (i frastødning) til dominerende alleler af et andet gen, der giver fordele for deres bærer (for eksempel, aB), der er forbundet med det "gode" tillader varigheden af det "dårlige".
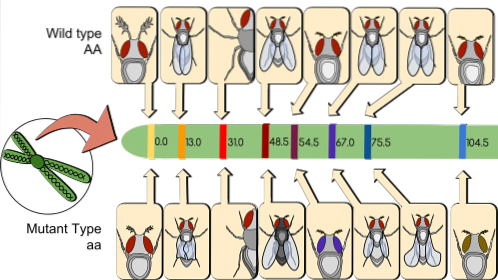
En vigtig konsekvens af kobling er, at det gør det muligt at bestemme afstanden mellem koblede gener. Dette viste sig at være historisk sandt og førte til dannelsen af de første genetiske kort..
Til dette var det nødvendigt at forstå, at homologe kromosomer kan krydse hinanden under meiose i en proces kaldet rekombination..
Ved rekombination produceres forskellige gameter til dem, som et individ kun kunne producere ved segregering. Da rekombinanter kan tælles, er det muligt matematisk at udtrykke, hvor langt et gen er adskilt fra et andet.
I koblings- og rekombinationskort tælles individer, der er rekombinante mellem et bestemt genpar. Derefter beregnes dens procent i forhold til den samlede beskæftigede kortlægningspopulation..
Efter konvention er en procent (1%) rekombination en genetisk kortenhed (umg). For eksempel findes i en kortlægningspopulation på 1000 individer 200 rekombinanter blandt de genetiske markører. TIL/til Y B/b. Derfor er afstanden, der adskiller dem på kromosomet, 20 umg.
I øjeblikket kaldes 1 umg (hvilket er 1% rekombination) cM (centi Morgan). I ovenstående tilfælde er afstanden mellem TIL/til Y B/b er 20 cM.
I et genetisk kort kan der tilføjes afstande i cM, men åbenlyst kan rekombinationsprocenter ikke tilføjes. Gener skal altid kortlægges, der er langt nok fra hinanden til at kunne måle korte afstande.
Hvis afstanden mellem to markører er meget høj, er sandsynligheden for, at der er en rekombinationshændelse mellem dem, lig med 1. Derfor vil de altid rekombineres, og disse gener vil opføre sig som om de var uafhængigt fordelt, selvom de er forbundet..
På den anden side er kortene målt i cM af forskellige årsager ikke lineært relateret til mængden af involveret DNA. Desuden er mængden af DNA pr. CM ikke universel, og for hver bestemt art er det en særlig og gennemsnitlig værdi..



Endnu ingen kommentarer