Det cellekryds de er de kontaktbroer, der findes mellem de cytoplasmatiske membraner mellem tilstødende celler eller mellem en celle og matrixen. Krydsene afhænger af den undersøgte vævstype og fremhæver de eksisterende forbindelser mellem epitel-, muskel- og nerveceller.
I celler er der molekyler relateret til adhæsion mellem dem. Imidlertid er der behov for yderligere elementer for at øge stabiliteten af bindingen i væv. Dette opnås med cellekryds.
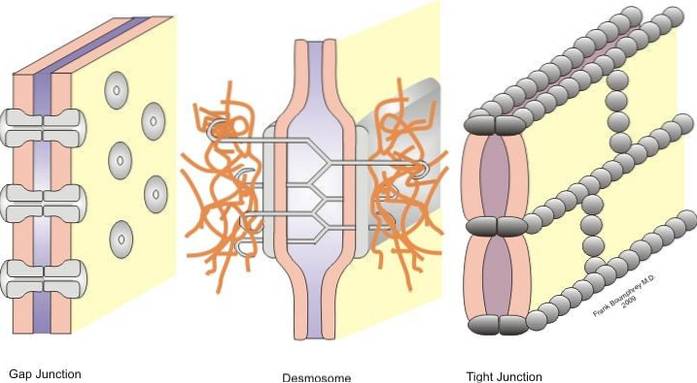
Kryds klassificeres i symmetriske kryds (tæt kryds, bælte desmosomer og spaltekryds) og asymmetriske kryds (hemidesmosomer).
Tætte kryds, bælte desmosomer, punkt desmosomer og hemidesmosomer er kryds, der muliggør forankring; mens kløftkrydsningerne opfører sig som foreningsbroer mellem naboceller, hvilket tillader udveksling af opløste stoffer mellem cytoplasmerne.
Bevægelsen af opløste stoffer, vand og ioner sker gennem og mellem individuelle cellulære komponenter. Således er der den transcellulære vej, der styres af en række kanaler og transportører. I modsætning til den paracellulære vej, som reguleres af kontakter mellem celler - det vil sige cellekryds.
I planter finder vi cellekryds, der ligner kløftkryds, kaldet plasmodesmata. Selvom de adskiller sig i struktur, er funktionen den samme.
Fra et medicinsk synspunkt oversættes visse mangler i celleforbindelser til erhvervede eller nedarvede sygdomme forårsaget af skade på epitelbarrieren.
Artikelindeks
Levende organismer består af diskrete og varierede strukturer kaldet celler. Disse afgrænses af en plasmamembran, der holder dem adskilt fra det ekstracellulære miljø..
Men selvom de er komponenterne i levende væsener, ligner de ikke mursten, da de ikke er isoleret fra hinanden.
Celler er elementer, der er i kommunikation med hinanden og med det ekstracellulære miljø. Derfor skal der være en måde, hvorpå celler kan danne væv og kommunikere, mens membranen forbliver intakt..
Dette problem kan løses takket være tilstedeværelsen af cellekryds, der findes i epithelia. Disse fagforeninger dannes mellem to tilstødende celler klassificeres i henhold til hver enkelt funktion i symmetriske og asymmetriske fagforeninger.
Hemidesmosomes hører til asymmetriske fagforeninger, og stramme fagforeninger, bånddesmosomer, desmosomer og kløftforeninger til symmetriske fagforeninger. Nedenfor beskriver vi hver fagforening i detaljer.
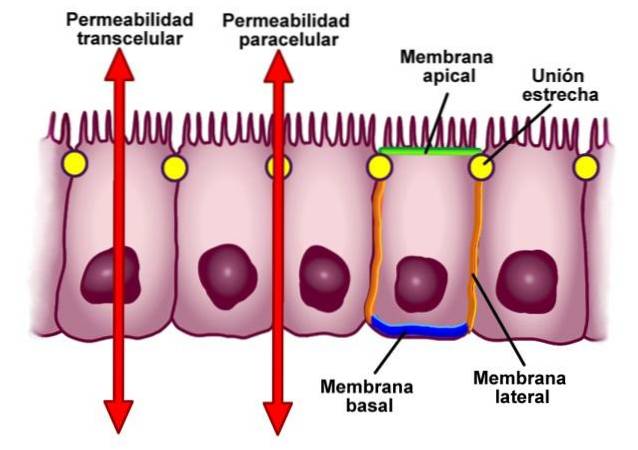
Stramme kryds, også kendt i litteraturen som okklusive kryds, er sektorer i cellemembranerne i naboceller, der er tæt forbundet - som navnet "tæt kryds" indikerer.
Under gennemsnitlige betingelser adskilles cellerne med en afstand på 10-20 nm. I tilfælde af tæt kryds reduceres denne afstand imidlertid betydeligt, og membranerne i begge celler fører til berøring eller endda fusion..
Et typisk tæt kryds er placeret mellem sidevæggene i nabocellerne i en minimumsafstand fra deres apikale overflader..
I epitelvæv laver alle celler sådanne kryds for at forblive sammen. I denne interaktion er celler arrangeret i et mønster, der minder om en ring. Disse fagforeninger dækker hele omkredsen.
De stramme kontaktregioner omgiver hele celleoverfladen. Disse regioner danner anastomiserede kontaktstrimler af transmembrane proteiner kendt som occludin og claudin. Begrebet anastomose henviser til foreningen af visse anatomiske elementer.
Disse to proteiner tilhører gruppen af tetraespaniner. De er kendetegnet ved at have fire transmembrane domæner, to ydre sløjfer og to relativt korte cytoplasmatiske haler..
Occludin har vist sig at interagere med fire andre proteinmolekyler, kaldet zonule occludin og forkortet som ZO. Denne sidste gruppe inkluderer proteinerne Z01, ZO2, ZO3 og afadin.
Claudin er på den anden side en familie på 16 proteiner, der udgør en række lineære fibriller i tætte kryds, som gør det muligt for denne krydsning at påtage sig rollen som en "barriere" i den paracellulære vej..
Nektiner og forbindelsesadhæsionsmolekyler (JAM) vises også i tætte kryds. Disse to molekyler findes som homodimerer i det intracellulære rum..
Nektiner er forbundet med actinfilamenter gennem proteinet afadin. Sidstnævnte synes at være afgørende, da de i sletningerne af genet, der koder for afadin hos gnavere, fører til embryonets død.
Disse typer forbindelser mellem celler udfører to vigtige funktioner. Den første er at bestemme polariteten af cellerne i epitelet, adskille det apikale domæne fra det basolaterale domæne og forhindre unødig diffusion af lipider, proteiner og andre biomolekyler i at finde sted..
Som vi nævnte i definitionen er epitelcellerne grupperet i en ring. Denne struktur adskiller den apikale overflade af cellen fra den laterale og den basale, hvilket etablerer differentieringen mellem domænerne.
Denne adskillelse betragtes som et af de vigtigste begreber i studiet af fysiologi af epitel..
For det andet forhindrer stramme krydsninger den frie passage af stoffer gennem epitelcellelaget, hvilket oversættes til en barriere for den paracellulære vej..
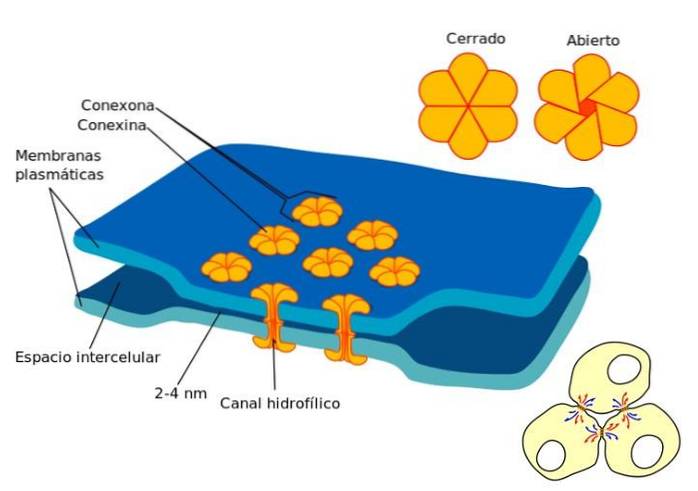
Mellemrumsknuderne findes i regioner uden begrænsende cytoplasmisk membran mellem naboceller. I en kløftkryds forbinder cellernes cytoplasmer, og der oprettes en fysisk forbindelse, hvor passage af små molekyler kan forekomme..
Denne klasse af kryds findes i næsten alle epithelia og i andre typer væv, hvor de tjener en række forskellige formål..
For eksempel kan spaltede kryds i forskellige væv åbne eller lukke som reaktion på ekstracellulære signaler, som det er tilfældet med neurotransmitteren dopamin. Tilstedeværelsen af dette molekyle reducerer kommunikationen mellem en klasse af neuroner i nethinden som reaktion på øget lysintensitet..
Spaltskæringspunkterne består af proteiner kaldet connexins. Således opnås en "connexon" ved foreningen af seks connexin-monomerer. Denne struktur er en hul cylinder, der findes på tværs af den cytoplasmatiske membran.
Forbindelserne er arrangeret på en sådan måde, at der dannes en ledning mellem cytoplasmaerne i tilstødende celler. Også forbindelserne har tendens til at aggregeres og danne en slags plader..
Takket være dannelsen af disse knudepunkter kan visse molekylers bevægelse mellem naboceller forekomme. Størrelsen på molekylet, der skal transporteres, er afgørende, den optimale diameter er 1,2, ligesom calciumioner og cyklisk adenosinmonophosphat.
Specifikt er de uorganiske ioner og vandopløselige molekyler, der kan overføres fra en cellecytoplasma til det tilstødende cytoplasma..
Calciumkoncentrationer spiller en afgørende rolle i denne kanal. Når calciumkoncentrationen stiger, har de aksiale kanaler en tendens til at lukke.
På denne måde deltager kløftkryds aktivt i den elektriske og kemiske koblingsproces mellem celler, som det sker i hjertets muskelceller, som er ansvarlige for transmission af elektriske impulser..
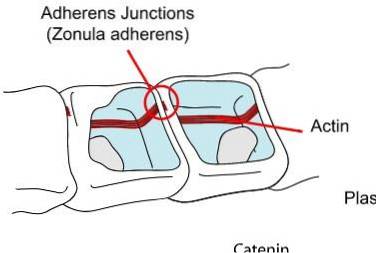
Under de tætte samlinger finder vi ankerledene. Generelt er disse placeret nær den apikale overflade af epitelet. I denne gruppe kan vi skelne mellem tre hovedgrupper, zonula adherens eller belt desmosome, macula adherens eller punktlig desmosome og desmosome.
I denne type krydsning adskilles de tilstødende cellemembraner, der er forbundet med zonulaer og adhærente makulaer, med en relativt stor celleafstand - sammenlignet med det minimale rum, der findes i tilfælde af tætte kryds..
Det intercellulære rum er optaget af proteiner, der hører til familien af cadheriner, desmogleiner og desmocholiner, der er knyttet til cytoplasmiske plaques, der præsenterer andre proteiner kaldet desmoplakin, placoglobin og placophilin.
Som i tilfælde af tætte samlinger observerer vi også i ankerledene mønsteret for arrangementet i form af en ring eller bælte. Zonula-adhærenserne er associeret med actinmikrofilm gennem interaktion mellem to proteiner: cadheriner og cateniner..
I nogle tilfælde er denne struktur kendt som et desmosom, det er en punktformet union, der er forbundet med mellemliggende filamenter dannet af keratin. I denne sammenhæng kaldes disse keratinstrukturer "tonofilimanetos". Filamenterne strækker sig fra punkt til punkt i epitelceller.
Disse giver styrke og stivhed til epitelcellerne. Det antages således, at dets hovedfunktion er relateret til styrkelse og stabilisering af tilstødende celler..
Desmosomer kan sammenlignes med en slags nitte eller svejsning, da de ligner separate små prikker og ikke kontinuerlige bånd.
Vi finder denne type knudepunkt i de indskudte skiver, der forbinder kardiocytterne i hjertemusklen og i hjernehinderne, der strækker den ydre overflade af hjernen og rygmarven..
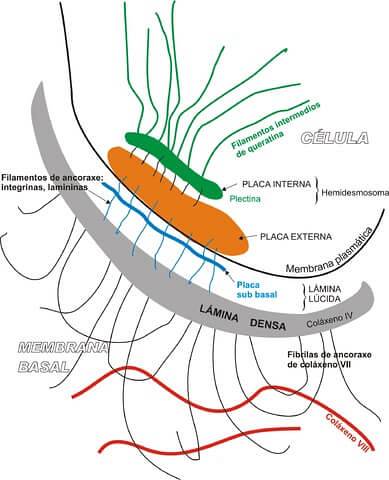
Hemidesmosomes falder ind under kategorien asymmetriske kryds. Disse strukturer har den funktion at forankre epitelcellens basale domæne med den underliggende basalaminat.
Udtrykket hemidesmosome bruges, fordi denne struktur vises bogstaveligt "halv" desmosom. Men set fra deres biokemiske sammensætning er begge fagforeninger helt forskellige..
Det er vigtigt at præcisere, at desmosomer er ansvarlige for at klæbe en nabocelle til en anden, mens hemidesmosomets funktion er at forene cellen med basal lamina.
I modsætning til makula-adhærenterne eller desmosomet har hemidesmosomer en anden struktur, der består af: en cytoplasmatisk laminat associeret med mellemliggende filamenter og en plade af eksterne membraner, som er ansvarlig for at forbinde hemidesmosomet med basalaminen ved hjælp af et ankerfilament.
En af funktionerne med hemidesmosomer er at øge den globale stabilitet af epitelvæv takket være tilstedeværelsen af mellemliggende cytoskeletale filamenter fastgjort til komponenterne i basal lamina.
Planteriget mangler de fleste af de ovenfor beskrevne cellekryds, med undtagelse af en funktionel modstykke, der minder om kløftkryds..
I planter er cytoplasmerne i tilstødende celler forbundet via veje eller kanaler kaldet plasmodesmata..
Denne struktur skaber et kontinuum fra en plantecelle til den næste. Selv om det adskiller sig strukturelt fra kløftkryds, har de meget ens roller, der tillader passage af små ioner og molekyler..
Fra et medicinsk synspunkt er celleforbindelser et relevant emne. Mutationer i generne, der koder for de proteiner, der er involveret i krydsene, har vist sig at oversætte til kliniske patologier.
For eksempel, hvis der er en bestemt mutation i genet, der koder for en bestemt type claudin (et af proteinerne, der formidler interaktionen ved tætte kryds), forårsager det en sjælden sygdom hos mennesker..
Dette er nedsat magnesiumtabsyndrom, og symptomer inkluderer lavt magnesium og anfald..
Derudover har en mutation i genet, der koder for nectin 1-proteinet, vist sig at være ansvarlig for kløftesyndrom. Denne tilstand betragtes som en af de mest almindelige misdannelser hos nyfødte..
Mutationer i nektin 1-genet er også blevet forbundet med en anden tilstand kaldet ektoparisk dysplasi, der påvirker menneskelig hud, hår, negle og tænder..
Pemphigus foliaceus er en blærende hudsygdom bestemt af autoantistoffer mod desmoglein 1, et nøgleelement, der er ansvarlig for at opretholde sammenhængen i epidermis.



Endnu ingen kommentarer